Aceștia sunt compuși organici cu molecul mare, biopolimeri, construiți din 20 de tipuri de reziduuri de L-p-aminoacizi conectate într-o anumită secvență în lanțuri lungi. Greutatea moleculară a proteinelor variază de la 5 mii la 1 milion. Numele „alb” a fost dat pentru prima dată substanței ouălor de păsări, care se coagulează atunci când este încălzită într-o masă albă insolubilă. Termenul a fost extins ulterior la alte substanțe cu proprietăți similare izolate de la animale și plante.
Orez. 1. Cei mai complexi biopolimeri sunt proteinele. Macromoleculele lor constau din monomeri, care sunt aminoacizi. Fiecare aminoacid are două grupe funcționale: o grupă carboxil și o grupă amino. Toată diversitatea proteinelor este creată ca urmare a diferitelor combinații de 20 de aminoacizi.
Proteinele predomină peste toți ceilalți compuși prezenți în organismele vii, de obicei reprezentând mai mult de jumătate din greutatea lor uscată. Se presupune că există câteva miliarde de proteine individuale în natură (de exemplu, mai mult de 3 mii de proteine diferite sunt prezente numai în bacteria E. coli).
Proteinele joacă un rol cheie în procesele de viață ale oricărui organism. Proteinele includ enzime, cu participarea cărora au loc toate transformările chimice în celulă (metabolism); controlează acțiunea genelor; cu participarea lor, se realizează acțiunea hormonilor, se efectuează transportul transmembranar, inclusiv generarea de impulsuri nervoase. Ele sunt parte integrantă a sistemului imunitar (imunoglobuline) și a sistemului de coagulare, formează baza osului și a țesutului conjunctiv și sunt implicate în transformarea și utilizarea energiei.
Istoria cercetării proteinelor
Primele încercări de a izola proteinele au fost făcute încă din secolul al XVIII-lea. La începutul secolului al XIX-lea au apărut primele lucrări privind studiul chimic al proteinelor. Oamenii de știință francezi Joseph Louis Gay-Lussac și Louis Jacques Thénard au încercat să stabilească compoziția elementară a proteinelor din diferite surse, ceea ce a marcat începutul unor studii analitice sistematice, datorită cărora s-a ajuns la concluzia că toate proteinele sunt similare în setul de elemente incluse în alcătuirea lor. În 1836, chimistul olandez G. J. Mulder a propus prima teorie a structurii substanțelor proteice, conform căreia toate proteinele au un anumit radical ipotetic (C 40 H 62 N 10 O 12), asociat în diverse proporții cu atomi de sulf și fosfor. El a numit acest radical „proteină” (de la proteina greacă - în primul rând, principal). Teoria lui Mulder a contribuit la creșterea interesului pentru studiul proteinelor și la îmbunătățirea metodelor de chimie a proteinelor. Au fost dezvoltate tehnici de izolare a proteinelor prin extracție cu soluții de săruri neutre, iar proteinele au fost obținute pentru prima dată sub formă cristalină (unele proteine vegetale). Pentru a analiza proteinele, au început să-și folosească digestia preliminară cu acizi și alcalii.
În același timp, a început să se acorde o atenție sporită studiului funcției proteinelor. Jens Jakob Berzelius a fost primul care a sugerat în 1835 că aceștia joacă rolul de biocatalizatori. Curând, au fost descoperite enzime proteolitice - pepsina (T. Schwann, 1836) și tripsina (L. Corvisart, 1856), care au atras atenția asupra fiziologiei digestiei și asupra analizei produselor formate în timpul descompunerii nutrienților. Studiile ulterioare ale structurii proteinelor și lucrările asupra sintezei chimice a peptidelor au dus la apariția ipotezei peptidelor, conform căreia toate proteinele sunt construite din aminoacizi. Până la sfârșitul secolului al XIX-lea, majoritatea aminoacizilor care alcătuiesc proteinele au fost studiați.
La începutul secolului al XX-lea, chimistul german Emil Hermann Fischer a fost primul care a folosit metodele chimiei organice pentru a studia proteinele și a demonstrat că proteinele constau din β-aminoacizi legați între ei printr-o legătură amidă (peptidică). Mai târziu, datorită utilizării metodelor fizico-chimice de analiză, a fost determinată masa moleculară a multor proteine, a fost stabilită forma sferică a proteinelor globulare, a fost efectuată analiza de difracție cu raze X a aminoacizilor și peptidelor și s-au efectuat metode de analiză cromatografică. dezvoltat (vezi cromatografia).
Primul hormon proteic a fost izolat (Frederick Grant Banting, John James Rickard McLeod, 1922), a fost dovedită prezența gammaglobulinelor în anticorpi și a fost descrisă funcția enzimatică a miozinei proteinei musculare (Vladimir Aleksandrovich Engelhardt, M. N. Lyubimova, 1939) . Pentru prima dată s-au obţinut enzime sub formă cristalină - urază (J.B. Saliner, 1926), pepsină (J.H. Nortron, 1929), lizozimă (E.P. Abraham, Robert Robinson, 1937).

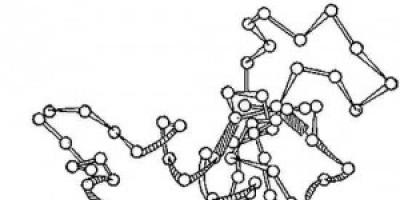
Orez. 2. Schema structurii tridimensionale a enzimei lizozime. Cercuri - aminoacizi; fire - legături peptidice; dreptunghiurile umbrite sunt legături disulfurice. Sunt vizibile secțiuni spiralizate și alungite ale lanțului polipeptidic.
În anii 1950 s-a dovedit organizarea pe trei niveluri a moleculelor de proteine - prezența unei structuri primare, secundare și terțiare; a creat un analizor automat de aminoacizi (Stanford Moore, William Howard Stein, 1950). În anii 60 s-au încercat sintetizarea chimică a proteinelor (insulina, ribonuclează). Metodele de analiză prin difracție cu raze X au fost îmbunătățite semnificativ; a fost creat un dispozitiv - un secvențior (P. Edman, G. Begg, 1967), care a făcut posibilă determinarea secvenței de aminoacizi dintr-un lanț polipeptidic. Consecința acestui lucru a fost stabilirea structurii a câteva sute de proteine dintr-o varietate de surse. Printre acestea se numără enzimele proteolitice (pepsină, tripsină, chimotripsină, subtilizină, carboxipeptidaze), mioglobine, hemoglobine, citocromi, lizozime, imunoglobuline, histone, neurotoxine, proteine ale anvelopei virale, hormoni proteino-peptidici. Ca urmare, au apărut premisele pentru rezolvarea problemelor presante din enzimologie, imunologie, endocrinologie și alte domenii ale chimiei biologice.
La sfârșitul secolului al XX-lea, s-au înregistrat progrese semnificative în studierea rolului proteinelor în sinteza matriceală a biopolimerilor, înțelegerea mecanismelor de acțiune a acestora în diferite procese de viață ale organismelor și stabilirea legăturii dintre structura și funcția acestora. Îmbunătățirea metodelor de cercetare și apariția unor noi metode de separare a proteinelor și peptidelor au fost de mare importanță.
Dezvoltarea unei metode eficiente de analiză a secvenței de nucleotide din acizii nucleici a făcut posibilă simplificarea și accelerarea semnificativă a determinării secvenței de aminoacizi din proteine. Acest lucru s-a dovedit a fi posibil deoarece ordinea aminoacizilor dintr-o proteină este determinată de secvența nucleotidelor din gena care codifică această proteină (fragment). În consecință, cunoscând aranjamentul nucleotidelor din această genă și codul genetic, se poate prezice cu exactitate în ce ordine se află aminoacizii în lanțul polipeptidic al unei proteine. Odată cu progresele în analiza structurală a proteinelor, s-au obținut rezultate semnificative în studiul organizării lor spațiale, mecanismelor de formare și acțiune a complexelor supramoleculare, inclusiv ribozomi și alte organite celulare, cromatina, virusuri etc.
Structura proteinei
Aproape toate proteinele sunt construite din 20 de α-aminoacizi aparținând seriei L și sunt aceleași în aproape toate organismele. Aminoacizii din proteine sunt legați între ei printr-o legătură peptidică -CO-NH-, care este formată din grupa carboxil și -amino a resturilor de aminoacizi vecine: doi aminoacizi formează o dipeptidă în care carboxilul terminal (-COOH) şi gruparea amino (H2N-) rămân libere, la care se pot adăuga noi aminoacizi pentru a forma un lanţ polipeptidic.
Secțiunea lanțului pe care se află grupa terminală H 2 N se numește N-terminal, iar partea opusă acestuia se numește C-terminal. Varietatea uriașă de proteine este determinată de secvența de aranjare și de numărul de reziduuri de aminoacizi pe care le conțin. Deși nu există o distincție clară, lanțurile scurte sunt de obicei numite peptide sau oligopeptide (din oligo...), iar polipeptidele (proteinele) sunt de obicei înțelese ca lanțuri formate din 50 sau mai multe. Cele mai comune proteine sunt cele care conțin 100-400 de resturi de aminoacizi, dar există și cele ale căror molecule sunt formate din 1000 sau mai multe reziduuri. Proteinele pot consta din mai multe lanțuri polipeptidice. În astfel de proteine, fiecare lanț polipeptidic este numit subunitate.
Structura spațială a proteinelor

Orez. 3. Proteinele din toate organismele sunt formate din 20 de tipuri de aminoacizi. Fiecare proteină este caracterizată de un anumit sortiment și raport cantitativ de aminoacizi. În moleculele proteice, aminoacizii sunt legați între ei prin legături peptidice (- CO - NH -) într-o secvență liniară, constituind așa-numita structură primară a proteinei. Linia superioară - aminoacizi liberi cu grupe laterale R1, R2, R3; concluzie - aminoacizii sunt legați prin legături peptidice.
Lanțul polipeptidic este capabil să formeze și să mențină în mod spontan o structură spațială specială. Pe baza formei moleculelor de proteine, proteinele sunt împărțite în fibrilare și globulare. În proteinele globulare, unul sau mai multe lanțuri polipeptidice sunt pliate într-o structură sferică compactă sau globule. De obicei, aceste proteine sunt foarte solubile în apă. Acestea includ aproape toate enzimele, proteinele de transport al sângelui și multe proteine de depozitare. Proteinele fibrilare sunt molecule sub formă de fire ținute împreună prin legături încrucișate și formează fibre lungi sau structuri stratificate. Au o rezistență mecanică ridicată, sunt insolubile în apă și îndeplinesc în principal funcții structurale și de protecție. Reprezentanții tipici ai acestor proteine sunt cheratinele de păr și lână, fibroina de mătase și colagenul de tendon.
Ordinea aminoacizilor legați covalent dintr-un lanț polipeptidic este numită secvența de aminoacizi sau structura primară a proteinelor. Structura primară a fiecărei proteine, codificată de gena corespunzătoare, este constantă și poartă toate informațiile necesare pentru formarea structurilor de nivel superior. Numărul potențial de proteine care se pot forma din 20 de aminoacizi este practic nelimitat.
Ca rezultat al interacțiunii grupurilor laterale de reziduuri de aminoacizi, secțiunile individuale relativ mici ale lanțului polipeptidic iau una sau alta conformație (tip de pliere), cunoscută ca structura secundară a proteinelor. Elementele sale cele mai caracteristice sunt α-helix și β-structură care se repetă periodic. Structura secundară este foarte stabilă. Deoarece este determinată în mare măsură de secvența de aminoacizi a regiunii proteice corespunzătoare, devine posibil să o prezicăm cu un anumit grad de probabilitate. Termenul „a-helix” a fost introdus de biochimistul, fizicianul și chimistul american Linus Carl Pauling, care a descris aranjarea lanțului polipeptidic în proteina a-keratina sub forma unei spirale drepte (a-helix poate fi comparat cu un cablu telefonic). Pentru fiecare tură a unui astfel de helix într-o proteină există 3,6 reziduuri de aminoacizi. Aceasta înseamnă că gruparea -C=O a unei legături peptidice formează o legătură de hidrogen cu gruparea -NH a altei legături peptidice, patru resturi de aminoacizi la distanță de prima. În medie, fiecare regiune α-helical include până la 15 aminoacizi, ceea ce corespunde la 3-4 spire ale helixului. Dar în fiecare proteină individuală, lungimea helixului poate diferi foarte mult de această valoare. În secțiune transversală, α-helixul are forma unui disc, din care lanțurile laterale de aminoacizi sunt îndreptate spre exterior.
Structura, sau? -stratul pliat, poate fi format din mai multe secțiuni ale lanțului polipeptidic. Aceste secțiuni sunt întinse și așezate paralel unele cu altele, conectate între ele prin legături de hidrogen care apar între legăturile peptidice. Ele pot fi orientate în direcții identice sau opuse (direcția de mișcare de-a lungul lanțului polipeptidic este de obicei considerată a fi de la capătul N-terminal la capătul C-terminal). În primul caz, stratul pliat se numește paralel, în al doilea - antiparalel. Acesta din urmă se formează atunci când lanțul peptidic face o întoarcere bruscă înapoi, formând o îndoire (a-bend). Lanțurile laterale de aminoacizi sunt orientate perpendicular pe plan? -strat.
Conținut relativ? -secţiuni în spirală şi? -structurile pot varia mult între diferitele proteine. Există proteine cu predominanța α-helix (aproximativ 75% din aminoacizi în mioglobină și hemoglobină), iar principalul tip de pliere a lanțului în multe proteine fibrilare (inclusiv fibroina de mătase, β-keratina) este α-helix. -structura. Regiunile lanțului polipeptidic care nu pot fi clasificate în niciuna dintre conformațiile descrise mai sus se numesc bucle de conectare. Structura lor este determinată în principal de interacțiunile dintre lanțurile laterale ale aminoacizilor, iar în molecula oricărei proteine se încadrează într-un mod strict definit.
Structura terțiară se numește structura spațială a proteinelor globulare. Dar adesea acest concept se referă la metoda de pliere a lanțului polipeptidic în spațiu, caracteristică fiecărei proteine specifice. Structura terțiară este formată din lanțul polipeptidic al unei proteine în mod spontan, aparent, de-a lungul unei anumite căi(e) de coagulare cu formarea preliminară a elementelor de structură secundară. Dacă stabilitatea structurii secundare se datorează legăturilor de hidrogen, atunci structura terțiară este fixată de un sistem divers de interacțiuni necovalente: hidrogen, interacțiuni ionice, intermoleculare, precum și contacte hidrofobe între lanțurile laterale de amino nepolar. reziduuri acide.
În unele proteine, structura terțiară este în continuare stabilizată prin formarea de legături disulfurice (legături -S-S-) între reziduurile de cisteină. De regulă, în interiorul globului proteic există lanțuri laterale de aminoacizi hidrofobi asamblate în miez (transferul lor în interiorul globului proteic este favorabil termodinamic), iar la periferie sunt reziduuri hidrofile și unele hidrofobe. Globulul proteic este înconjurat de câteva sute de molecule de apă de hidratare, care este necesară pentru stabilitatea moleculei proteice și este adesea implicată în funcționarea acesteia. Structura terțiară este mobilă, secțiunile sale individuale se pot deplasa, ceea ce duce la tranziții conformaționale care joacă un rol semnificativ în interacțiunea proteinei cu alte molecule.
Structura terțiară este baza proprietăților funcționale ale unei proteine. Determină formarea ansamblurilor de grupuri funcționale în proteină - centri activi și zone de legare, le conferă geometria necesară, permite crearea unui mediu intern, care este o condiție prealabilă pentru apariția multor reacții și asigură interacțiunea cu alte proteine. .
Structura terțiară a proteinelor corespunde clar structurii sale primare; probabil că există un cod stereochimic încă nedescifrat care determină natura plierii proteinelor. Cu toate acestea, una și aceeași metodă de aranjare spațială corespunde de obicei nu unei singure structuri primare, ci unei întregi familii de structuri în care doar o mică fracțiune (până la 20-30%) de reziduuri de aminoacizi poate coincide, dar în anumite cazuri. plasează în lanț asemănarea resturilor de aminoacizi se păstrează. Rezultatul este formarea unor familii mari de proteine caracterizate prin structură primară terțiară similară și mai mult sau mai puțin similară și, de regulă, funcție comună. Acestea sunt, de exemplu, proteine ale organismelor de diferite specii care au aceeași funcție și sunt înrudite evolutiv: mioglobine și hemoglobine, tripsina, chimotripsină, elastaza și alte proteinaze animale.

Orez. 4. Ca rezultat al combinației mai multor macromolecule proteice cu o structură terțiară, o structură proteică cuaternară se formează într-un complex complex. Un exemplu de astfel de proteine complexe este hemoglobina, constând din patru macromolecule.
Adesea, mai ales la proteinele mari, plierea unui lanț polipeptidic are loc prin formarea de către secțiuni individuale ale lanțului a unor elemente mai mult sau mai puțin autonome de structură spațială - domenii care pot avea autonomie funcțională, fiind responsabile de una sau alta activitate biologică a proteină. Astfel, domeniile N-terminale ale proteinelor de coagulare a sângelui asigură atașarea acestora la membrana celulară.
Există multe proteine ale căror molecule sunt un ansamblu de globule (subunități) ținute împreună prin interacțiuni hidrofobe, hidrogen sau legături ionice. Astfel de complexe se numesc proteine oligomerice, multimerice sau subunități. Dispunerea subunităților într-un complex proteic activ funcțional se numește structura cuaternară a proteinei. Unele proteine sunt capabile să formeze structuri de ordin superior, de exemplu, complexe multienzimatice, structuri extinse (proteine de acoperire bacteriofage), complexe supramoleculare care funcționează ca un întreg (de exemplu, ribozomi sau componente ale lanțului respirator mitocondrial).
Structura cuaternară permite crearea de molecule cu geometrii neobișnuite. Astfel, feritina, formată din 24 de subunități, are o cavitate internă, datorită căreia proteina reușește să lege până la 3000 de ioni de fier. În plus, structura cuaternară permite îndeplinirea mai multor funcții diferite într-o moleculă. Triptofan sintetaza combină enzimele responsabile pentru mai multe etape succesive ale sintezei aminoacidului triptofan.
Metode pentru studiul structurii proteinelor
Structura primară a proteinelor determină toate celelalte niveluri de organizare a moleculei proteice. Prin urmare, atunci când se studiază funcția biologică a diferitelor proteine, cunoașterea acestei structuri este importantă. Prima proteină pentru care a fost stabilită secvența de aminoacizi a fost hormonul pancreatic, insulina. Această lucrare, care a durat 11 ani, a fost realizată de biochimistul englez Frederick Sanger (1954). El a determinat localizarea a 51 de aminoacizi în molecula de hormon și a arătat că aceasta constă din 2 lanțuri legate prin legături disulfurice. Mai târziu, cea mai mare parte a lucrărilor de stabilire a structurii primare a proteinelor a fost automatizată.
Odată cu dezvoltarea metodelor de inginerie genetică, a devenit posibilă accelerarea în continuare a acestui proces prin determinarea structurii primare a proteinelor în conformitate cu rezultatele analizei secvenței de nucleotide din genele care codifică aceste proteine. Structura secundară și terțiară a proteinelor este studiată folosind metode fizice destul de complexe, de exemplu, dicroismul circular sau analiza de difracție de raze X a cristalelor de proteine. Structura terțiară a fost stabilită pentru prima dată de biochimistul englez John Cowdery Kendrew (1957) pentru proteina musculară mioglobina.

Orez. 5. Modelul moleculei de mioglobină (configurația spațială a moleculei)
Denaturarea proteinelor
Legăturile relativ slabe responsabile pentru stabilizarea structurilor secundare, terțiare și cuaternare ale proteinei sunt ușor distruse, ceea ce este însoțit de o pierdere a activității sale biologice. Distrugerea structurii proteice originale (native), numită denaturare, are loc în prezența acizilor și bazelor, cu încălzire, modificări ale puterii ionice și alte influențe. De regulă, proteinele denaturate sunt slab sau deloc solubile în apă. Cu un efect pe termen scurt și eliminarea rapidă a factorilor de denaturare, renaturarea proteinelor este posibilă cu refacerea completă sau parțială a structurii originale și a proprietăților biologice.
Clasificarea proteinelor
Complexitatea structurii moleculelor de proteine și varietatea extremă a funcțiilor pe care le îndeplinesc fac dificilă crearea unei clasificări unificate și clare a acestora, deși încercările de a face acest lucru au fost făcute în mod repetat de la sfârșitul secolului al XIX-lea. Pe baza compoziției lor chimice, proteinele sunt împărțite în simple și complexe (uneori numite proteide). Moleculele primelor constau numai din aminoacizi. Pe lângă lanțul polipeptidic în sine, proteinele complexe conțin componente non-proteice reprezentate de carbohidrați (glicoproteine), lipide (lipoproteine), acizi nucleici (nucleoproteine), ioni metalici (metaloproteine), grupare fosfat (fosfoproteine), pigmenți (cromoproteine), etc.
În funcție de funcțiile pe care le îndeplinesc, se disting mai multe clase de proteine. Cea mai diversă și mai specializată clasă este formată din proteine cu funcție catalitică - enzime care au capacitatea de a accelera reacțiile chimice care apar în organismele vii. În această calitate, proteinele participă la toate procesele de sinteză și descompunere a diverșilor compuși în timpul metabolismului, la biosinteza proteinelor și acizilor nucleici, la reglarea dezvoltării și diferențierii celulelor. Proteinele de transport au capacitatea de a lega selectiv acizii grași, hormoni și alți compuși și ioni organici și anorganici și apoi le transportă cu curent în locația dorită (de exemplu, hemoglobina este implicată în transferul de oxigen din plămâni către toate celulele corpul). Proteinele de transport efectuează, de asemenea, transportul activ de ioni, lipide, zaharuri și aminoacizi prin membranele biologice.
Proteinele structurale îndeplinesc o funcție de susținere sau de protecție; ei participă la formarea scheletului celular. Cele mai frecvente dintre ele sunt colagenul țesutului conjunctiv, keratina, unghiile și pene, elastina celulelor vasculare și multe altele. În combinație cu lipidele, ele sunt baza structurală a membranelor celulare și intracelulare.
Un număr de proteine îndeplinesc o funcție de protecție. De exemplu, imunoglobulinele (anticorpii) vertebratelor, având capacitatea de a lega microorganismele și substanțele patogene străine, neutralizează efectele lor patogene asupra organismului și previn proliferarea celulară. Fibrinogenul și trombina sunt implicate în procesul de coagulare a sângelui. Multe substanțe proteice secretate de bacterii, precum și componente ale unor nevertebrate, sunt clasificate drept toxine.
Unele proteine (de reglare) sunt implicate în reglarea activității fiziologice a corpului în ansamblu, a organelor, celulelor sau proceselor individuale. Ei controlează transcripția genelor și sinteza proteinelor; acestea includ hormoni peptido-proteici secretați de glandele endocrine. Proteinele de depozitare a semințelor oferă nutrienți pentru etapele inițiale ale dezvoltării embrionului. Acestea includ și cazeina, albumina de albuș de ou (ovalbumină) și multe altele. Datorită proteinelor, celulele musculare dobândesc capacitatea de a se contracta și în cele din urmă asigură mișcarea corpului. Exemple de astfel de proteine contractile sunt actina musculară scheletică și miozina, precum și tubulina, care sunt componente ale cililor și flagelilor organismelor unicelulare; Ele asigură, de asemenea, divergența cromozomilor în timpul diviziunii celulare.
Proteinele receptorilor sunt ținta hormonilor și a altor compuși biologic activi. Cu ajutorul lor, celula percepe informații despre starea mediului extern. Ele joacă un rol important în transmiterea excitației nervoase și în mișcarea orientată a celulelor (chemotaxie). Transformarea și utilizarea energiei care intră în organism, precum și a energiei, au loc și cu participarea proteinelor sistemului bioenergetic (de exemplu, pigmentul vizual rodopsina, citocromii lanțului respirator). Există, de asemenea, multe proteine cu alte funcții, uneori destul de neobișnuite (de exemplu, plasma unor pești din Antarctica conține proteine care au proprietăți antigel).
Biosinteza proteinelor
Toate informațiile despre structura unei anumite proteine sunt „stocate” în genele corespunzătoare sub forma unei secvențe de nucleotide și sunt implementate în procesul de sinteză a șablonului. Mai întâi, informațiile sunt transferate (citite) de la molecula de ADN la ARN-ul mesager (ARNm) folosind enzima ARN polimeraza dependentă de ADN și apoi în ribozom de pe ARNm, ca pe o matrice în conformitate cu codul genetic, cu participarea a ARN-urilor de transport care livrează aminoacizi, are loc formarea lanțului polipeptidic.
Lanțurile polipeptidice sintetizate care ies din ribozom, pliându-se spontan, capătă caracteristica conformației proteinei și pot fi supuse modificării post-translaționale. Lanțurile laterale ale aminoacizilor individuali pot suferi modificări (hidroxilare, fosforilare etc.). De aceea, de exemplu, hidroxiprolina și hidroxilizina se găsesc în colagen (vezi). Modificarea poate fi, de asemenea, însoțită de ruperea legăturilor polipeptidice. În acest fel, de exemplu, are loc formarea unei molecule active de insulină, formată din două lanțuri legate prin legături disulfurice.

Orez. 6. Schema generală a biosintezei proteinelor.
Importanța proteinelor în nutriție
Proteinele sunt cele mai importante componente ale alimentelor animale și umane. Valoarea nutritivă a proteinelor este determinată de conținutul lor de aminoacizi esențiali, care nu sunt produși chiar în organism. În acest sens, proteinele vegetale sunt mai puțin valoroase decât proteinele animale: sunt mai sărace în lizină, metionină și triptofan și sunt mai greu de digerat în tractul gastrointestinal. Lipsa aminoacizilor esențiali din alimente duce la tulburări severe ale metabolismului azotului.
Proteinele sunt descompuse în aminoacizi liberi, care, după absorbția în intestin, intră și sunt distribuite în toate celulele. Unele dintre ele se descompun în compuși simpli cu eliberare de energie, utilizate pentru diferite nevoi de către celulă, iar altele merg la sinteza de noi proteine caracteristice unui anumit organism. (R. A. Matveeva, Enciclopedia Cyril și Methodius)
Numărarea proteinelor
- amiloid - amiloid;
- anionic - anionic;
- antivirus - antiviral;
- autoimună - autoimună;
- autolog - autologic;
- bacteriană - bacteriană;
- proteina Bence Jones;
- virus-induced - virus indus;
- viral - virus;
- viral nonstructural - virus nonstructural;
- viral structural - virus structural;
- virus-specific - specific virusului;
- greutate moleculară mare - greutate moleculară mare;
- conţinând hem - hem;
- heterolog - străin;
- hibrid - hibrid;
- glicozilat - glicat;
- globular - globular;
- denaturat - denaturat;
- care conțin fier - fier;
- galbenus - galbenus;
- proteina animală - proteină animală;
- protectiv - defensiv;
- imunitar - imunitar;
- imunogen - relevant din punct de vedere imunologic;
- legarea calciului;
- acru - acid;
- corpuscular - corpuscular;
- membrana - membrana;
- mielom - mielom;
- microzomal - microzomal;
- proteina din lapte - proteina din lapte;
- monoclonal - imunoglobulina monoclonal;
- proteină musculară - proteină musculară;
- nativ - nativ;
- nonhistone - nonhistone;
- defect - parțial;
- insolubil - insolubil;
- indigerabil - insolubil;
- non-enzimatică - nonenzimă;
- greutate moleculară mică - greutate moleculară mică;
- proteină nouă - proteină nouă;
- general - întreg;
- oncogen - oncoprotein;
- proteina de faza principala - anionica;
- proteina de faza acuta (inflamatie) - proteina de faza acuta;
- alimente - alimente;
- proteina plasmatică a sângelui - proteină plasmatică;
- placentar - placenta;
- uncoupling - decuplare;
- proteină a nervului regenerant;
- reglementare - reglementare;
- recombinare - recombinant;
- receptor - receptor;
- ribozomal - ribozomal;
- legare - legare;
- secretory protein - proteină secretorie;
- C-reactiv - C-reactiv;
- proteina din zer - proteina din zer, lactoproteina;
- tesut - tesut;
- toxic - toxic;
- himeric - himeric;
- întreg - întreg;
- citosol - citosol;
- proteina alcalina - proteina anionica;
- exogen - exogen;
- endogenă - proteină endogenă.
Citiți mai multe despre proteine în literatură:
- Volkenshtein M.V., Molecules and, M., 1965, cap. 3 - 5;
- Gaurowitz F., Chimia și funcțiile proteinelor, trans. din engleză, Moscova, 1965;
- Sissakyan N. M. și Gladilin K. L., Biochemical aspects of protein synthesis, în cartea: Advances in biological chemistry, vol. 7, M., 1965, p. 3;
- Stepanov V. M. Biologie moleculară. Structura și funcția proteinelor. M., 1996;
- Shamin A. N., Development of protein chemistry, M., 1966;
- Proteine și peptide. M., 1995-2000. T. 1-3;
- Biosinteza proteinelor și acizilor nucleici, ed. A. S. Spirina, M., 1965;
- Introducere în biologia moleculară, trad. din engleză, M., 1967
- Molecule și celule. [Sam. art.], trad. din engleză, M., 1966, p. 7 - 27, 94 - 106;
- Fundamentele biochimiei: Traducere din engleză M., 1981. T. 1;
- Problema proteinelor. M., 1995. T. 1-5;
- Proteinele. New York, 1975-79. 3 ed. V. 1-4.
Găsiți altceva interesant:
Veverițe- compuși organici cu greutate moleculară mare formați din reziduuri de α-aminoacizi.
ÎN compozitia proteinelor include carbon, hidrogen, azot, oxigen, sulf. Unele proteine formează complexe cu alte molecule care conțin fosfor, fier, zinc și cupru.
Proteinele au o greutate moleculară mare: albumina de ou - 36 000, hemoglobina - 152 000, miozina - 500 000. Pentru comparație: greutatea moleculară a alcoolului este de 46, acid acetic - 60, benzen - 78.
Compoziția de aminoacizi a proteinelor
Veverițe- polimeri neperiodici, ai căror monomeri sunt α-aminoacizi. De obicei, 20 de tipuri de α-aminoacizi sunt numite monomeri proteici, deși peste 170 dintre ei se găsesc în celule și țesuturi.
În funcție de dacă aminoacizii pot fi sintetizați în corpul oamenilor și al altor animale, aceștia se disting: aminoacizi neesențiali- poate fi sintetizat; aminoacizi esentiali- nu poate fi sintetizat. Aminoacizii esențiali trebuie să fie furnizați organismului prin alimente. Plantele sintetizează toate tipurile de aminoacizi.
În funcție de compoziția de aminoacizi, proteinele sunt: complete- conțin întregul set de aminoacizi; defect- unii aminoacizi lipsesc din compozitia lor. Dacă proteinele constau numai din aminoacizi, ele se numesc simplu. Dacă proteinele conțin, pe lângă aminoacizi, o componentă non-aminoacid (grup protetic), ele se numesc complex. Grupul protetic poate fi reprezentat de metale (metaloproteine), carbohidrați (glicoproteine), lipide (lipoproteine), acizi nucleici (nucleoproteine).
Toate aminoacizii conțin: 1) grupare carboxil (-COOH), 2) grupare amino (-NH 2), 3) radical sau grup R (restul moleculei). Structura radicalului este diferită pentru diferite tipuri de aminoacizi. În funcție de numărul de grupări amino și grupări carboxil incluse în compoziția aminoacizilor, acestea se disting: aminoacizi neutri având o grupare carboxil și o grupare amino; aminoacizi bazici având mai mult de o grupare amino; aminoacizi acizi având mai mult de o grupare carboxil.
Aminoacizii sunt compuși amfoteri, deoarece în soluție pot acționa atât ca acizi, cât și ca baze. În soluțiile apoase, aminoacizii există în diferite forme ionice.
Legătură peptidică
Peptide- substanțe organice formate din reziduuri de aminoacizi legate prin legături peptidice.
Formarea peptidelor are loc ca urmare a reacției de condensare a aminoacizilor. Când gruparea amino a unui aminoacid interacționează cu gruparea carboxil a altuia, între ele are loc o legătură covalentă azot-carbon, care se numește peptidă. În funcție de numărul de resturi de aminoacizi incluse în peptidă, există dipeptide, tripeptide, tetrapeptide etc. Formarea unei legături peptidice poate fi repetată de mai multe ori. Aceasta duce la formare polipeptide. La un capăt al peptidei există o grupare amino liberă (numită N-terminal), iar la celălalt există o grupare carboxil liberă (numită C-terminal).

Organizarea spațială a moleculelor proteice
Efectuarea anumitor funcții specifice de către proteine depinde de configurația spațială a moleculelor acestora; în plus, este nefavorabil din punct de vedere energetic pentru celulă să mențină proteinele într-o formă desfășurată, sub formă de lanț, prin urmare lanțurile polipeptidice sunt supuse plierii, dobândind un anumită structură tridimensională sau conformație. Sunt 4 niveluri organizarea spațială a proteinelor.
Structura primară a proteinei- secvența de aranjare a resturilor de aminoacizi în lanțul polipeptidic care alcătuiește molecula proteică. Legătura dintre aminoacizi este o legătură peptidică.

Dacă o moleculă de proteină este formată din doar 10 resturi de aminoacizi, atunci numărul de variante teoretic posibile ale moleculelor de proteine care diferă în ordinea alternanței aminoacizilor este de 10 20. Având 20 de aminoacizi, puteți face combinații și mai diverse din aceștia. În corpul uman au fost găsite aproximativ zece mii de proteine diferite, care diferă atât una de cealaltă, cât și de proteinele altor organisme.
Este structura primară a moleculei proteice care determină proprietățile moleculelor proteice și configurația sa spațială. Înlocuirea unui singur aminoacid cu altul într-un lanț polipeptidic duce la o schimbare a proprietăților și funcțiilor proteinei. De exemplu, înlocuirea celui de-al șaselea aminoacid glutamic cu valină în subunitatea β a hemoglobinei duce la faptul că molecula de hemoglobină în ansamblu nu își poate îndeplini funcția principală - transportul oxigenului; În astfel de cazuri, persoana dezvoltă o boală numită anemie cu celule falciforme.
Structura secundară- plierea ordonată a lanțului polipeptidic într-o spirală (pare un arc prelungit). Turnurile helixului sunt întărite de legăturile de hidrogen care apar între grupările carboxil și grupările amino. Aproape toate grupările CO și NH participă la formarea legăturilor de hidrogen. Ele sunt mai slabe decât cele peptidice, dar, repetate de multe ori, conferă stabilitate și rigiditate acestei configurații. La nivelul structurii secundare se regasesc proteine: fibroina (matase, panza de paianjen), keratina (par, unghii), colagen (tendoane).



Structura terțiară- împachetarea lanțurilor polipeptidice în globule, rezultată din formarea de legături chimice (hidrogen, ionice, disulfură) și stabilirea interacțiunilor hidrofobe între radicalii reziduurilor de aminoacizi. Rolul principal în formarea structurii terțiare îl joacă interacțiunile hidrofil-hidrofobe. În soluțiile apoase, radicalii hidrofobi tind să se ascundă de apă, grupându-se în interiorul globului, în timp ce radicalii hidrofili, ca urmare a hidratării (interacțiunii cu dipolii de apă), tind să apară la suprafața moleculei. În unele proteine, structura terțiară este stabilizată prin legături covalente disulfurice formate între atomii de sulf ai două reziduuri de cisteină. La nivelul structurii terțiare există enzime, anticorpi și unii hormoni.

Structura cuaternară caracteristic proteinelor complexe ale căror molecule sunt formate din două sau mai multe globule. Subunitățile sunt reținute în moleculă prin interacțiuni ionice, hidrofobe și electrostatice. Uneori, în timpul formării unei structuri cuaternare, între subunități apar legături disulfurice. Cea mai studiată proteină cu structură cuaternară este hemoglobină. Este format din două subunități α (141 de resturi de aminoacizi) și două subunități β (146 de resturi de aminoacizi). Cu fiecare subunitate este asociată o moleculă de hem care conține fier.
Dacă din anumite motive conformația spațială a proteinelor se abate de la normal, proteina nu își poate îndeplini funcțiile. De exemplu, cauza „boii vacii nebune” (encefalopatie spongiformă) este conformația anormală a prionilor, proteinele de suprafață ale celulelor nervoase.
Proprietățile proteinelor
Compoziția de aminoacizi și structura moleculei proteice o determină proprietăți. Proteinele combină proprietățile de bază și acide, determinate de radicalii aminoacizi: cu cât aminoacizii sunt mai acizi într-o proteină, cu atât sunt mai pronunțate proprietățile sale acide. Se determină capacitatea de a dona și de a adăuga H + proprietățile tampon ale proteinelor; Una dintre cele mai puternice soluții tampon este hemoglobina din celulele roșii din sânge, care menține pH-ul sângelui la un nivel constant. Există proteine solubile (fibrinogen), și există proteine insolubile care îndeplinesc funcții mecanice (fibroină, keratina, colagen). Sunt proteine active din punct de vedere chimic (enzime), sunt proteine inactive din punct de vedere chimic care sunt rezistente la diverse conditii de mediu si cele care sunt extrem de instabile.
Factori externi (caldura, radiatii ultraviolete, metale grele si sarurile acestora, modificari ale pH-ului, radiatii, deshidratare)
poate provoca perturbarea organizării structurale a moleculei proteice. Procesul de pierdere a conformației tridimensionale inerente unei molecule de proteină dată este numit denaturare. Cauza denaturarii este ruperea legaturilor care stabilizeaza o anumita structura proteica. Inițial, cele mai slabe legături sunt rupte, iar pe măsură ce condițiile devin mai stricte, se rup și cele mai puternice. Prin urmare, se pierd mai întâi structurile cuaternare, apoi cele terțiare și secundare. O modificare a configurației spațiale duce la o modificare a proprietăților proteinei și, ca urmare, face imposibilă ca proteina să își îndeplinească funcțiile biologice inerente. Dacă denaturarea nu este însoțită de distrugerea structurii primare, atunci poate fi reversibil, în acest caz, are loc auto-recuperarea conformației caracteristice proteinei. De exemplu, proteinele receptorilor membranari suferă o astfel de denaturare. Procesul de refacere a structurii proteinelor după denaturare se numește renaturare. Dacă restabilirea configurației spațiale a proteinei este imposibilă, atunci se numește denaturare ireversibil.
Funcțiile proteinelor
| Funcţie | Exemple și explicații |
|---|---|
| Constructie | Proteinele sunt implicate în formarea structurilor celulare și extracelulare: fac parte din membranele celulare (lipoproteine, glicoproteine), păr (keratina), tendoane (colagen) etc. |
| Transport | Hemoglobina proteică din sânge atașează oxigenul și îl transportă de la plămâni la toate țesuturile și organele, iar din acestea transferă dioxidul de carbon către plămâni; Compoziția membranelor celulare include proteine speciale care asigură transferul activ și strict selectiv al anumitor substanțe și ioni din celulă în mediul extern și înapoi. |
| de reglementare | Hormonii proteici iau parte la reglarea proceselor metabolice. De exemplu, hormonul insulina reglează nivelul de glucoză din sânge, promovează sinteza glicogenului și crește formarea grăsimilor din carbohidrați. |
| De protecţie | Ca răspuns la pătrunderea proteinelor străine sau a microorganismelor (antigene) în organism, se formează proteine speciale - anticorpi care le pot lega și neutraliza. Fibrina, formată din fibrinogen, ajută la oprirea sângerării. |
| Motor | Proteinele contractile actina și miozina asigură contracția musculară la animalele multicelulare. |
| Semnal | În membrana de suprafață a celulei sunt încorporate molecule de proteine care sunt capabile să-și schimbe structura terțiară ca răspuns la factorii de mediu, primind astfel semnale din mediul extern și transmitând comenzi către celulă. |
| Depozitare | În organismul animalelor, proteinele, de regulă, nu sunt stocate, cu excepția albuminei de ou și a cazeinei din lapte. Dar datorită proteinelor, unele substanțe pot fi stocate în organism; de exemplu, în timpul descompunerii hemoglobinei, fierul nu este îndepărtat din organism, ci este stocat, formând un complex cu proteina feritina. |
| Energie | Când 1 g de proteină se descompune în produse finite, se eliberează 17,6 kJ. În primul rând, proteinele se descompun în aminoacizi, apoi în produsele finale - apă, dioxid de carbon și amoniac. Cu toate acestea, proteinele sunt folosite ca sursă de energie numai atunci când alte surse (carbohidrați și grăsimi) sunt consumate. |
| catalitic | Una dintre cele mai importante funcții ale proteinelor. Furnizat de proteine - enzime care accelerează reacțiile biochimice care apar în celule. De exemplu, ribulozobifosfat carboxilaza catalizează fixarea CO 2 în timpul fotosintezei. |
Enzime
Enzime, sau enzime, sunt o clasă specială de proteine care sunt catalizatori biologici. Datorită enzimelor, reacțiile biochimice au loc cu o viteză extraordinară. Viteza reacțiilor enzimatice este de zeci de mii de ori (și uneori de milioane) mai mare decât viteza reacțiilor care au loc cu participarea catalizatorilor anorganici. Substanța asupra căreia acționează enzima se numește substrat.
Enzimele sunt proteine globulare, caracteristici structurale enzimele pot fi împărțite în două grupe: simple și complexe. Enzime simple sunt proteine simple, adică constau numai din aminoacizi. Enzime complexe sunt proteine complexe, adică În plus față de partea proteică, ele conțin un grup de natură non-proteică - cofactor. Unele enzime folosesc vitamine ca cofactori. Molecula de enzimă conține o parte specială numită centru activ. Centru activ- o mică secțiune a enzimei (de la trei până la douăsprezece reziduuri de aminoacizi), unde are loc legarea substratului sau substraturilor pentru a forma un complex enzimă-substrat. La terminarea reacției, complexul enzimă-substrat se descompune în enzimă și produs(i) de reacție. Unele enzime au (cu excepția activelor) centri alosterici- zone la care sunt atașați regulatorii de viteză a enzimelor ( enzime alosterice).

Reacțiile de cataliză enzimatică se caracterizează prin: 1) eficiență ridicată, 2) selectivitate strictă și direcție de acțiune, 3) specificitate substratului, 4) reglare fină și precisă. Substratul și specificitatea de reacție a reacțiilor de cataliză enzimatică sunt explicate prin ipotezele lui E. Fischer (1890) și D. Koshland (1959).
E. Fisher (ipoteza blocării tastelor) a sugerat că configurațiile spațiale ale situsului activ al enzimei și ale substratului trebuie să corespundă exact una cu cealaltă. Substratul este comparat cu „cheia”, enzima cu „lacătul”.
D. Koshland (ipoteza mână-mănuși) a sugerat că corespondența spațială dintre structura substratului și centrul activ al enzimei este creată numai în momentul interacțiunii lor unul cu celălalt. Această ipoteză se mai numește ipoteza corespondenței induse.
Viteza reacțiilor enzimatice depinde de: 1) temperatură, 2) concentrația enzimei, 3) concentrația substratului, 4) pH-ul. Trebuie subliniat faptul că, deoarece enzimele sunt proteine, activitatea lor este cea mai mare în condiții normale din punct de vedere fiziologic.
Majoritatea enzimelor pot funcționa doar la temperaturi cuprinse între 0 și 40°C. În aceste limite, viteza de reacție crește de aproximativ 2 ori la fiecare creștere de 10 °C a temperaturii. La temperaturi peste 40 °C, proteina este supusă denaturarii și activitatea enzimatică scade. La temperaturi apropiate de îngheț, enzimele sunt inactivate.
Pe măsură ce cantitatea de substrat crește, viteza reacției enzimatice crește până când numărul de molecule de substrat este egal cu numărul de molecule de enzimă. Cu o creștere suplimentară a cantității de substrat, viteza nu va crește, deoarece centrii activi ai enzimei sunt saturati. O creștere a concentrației de enzime duce la creșterea activității catalitice, deoarece un număr mai mare de molecule de substrat suferă transformări pe unitatea de timp.

Pentru fiecare enzimă, există o valoare optimă a pH-ului la care prezintă activitate maximă (pepsină - 2,0, amilaza salivară - 6,8, lipaza pancreatică - 9,0). La valori mai mari sau mai mici ale pH-ului, activitatea enzimatică scade. Odată cu schimbări bruște ale pH-ului, enzima se denaturează.
Viteza enzimelor alosterice este reglată de substanțe care se atașează de centrii alosterici. Dacă aceste substanțe accelerează o reacție, se numesc activatori, dacă încetinesc - inhibitori.
Clasificarea enzimelor
În funcție de tipul de transformări chimice pe care le catalizează, enzimele sunt împărțite în 6 clase:
- oxireductaze(transferul atomilor de hidrogen, oxigen sau electroni de la o substanță la alta - dehidrogenază),
- transferaze(transferul grupării metil, acil, fosfat sau amino de la o substanță la alta - transaminaza),
- hidrolaze(reacții de hidroliză în care din substrat se formează doi produși - amilază, lipază),
- liazele(adăugarea nehidrolitică la substrat sau detașarea unui grup de atomi din acesta, caz în care legăturile C-C, C-N, C-O, C-S pot fi rupte - decarboxilază),
- izomeraze(rearanjare intramoleculară - izomerază),
- ligaze(conectarea a două molecule ca urmare a formării legăturilor C-C, C-N, C-O, C-S - sintetaza).
Clasele sunt la rândul lor subdivizate în subclase și subsubclase. În clasificarea internațională actuală, fiecare enzimă are un cod specific, format din patru numere separate prin puncte. Primul număr este clasa, al doilea este subclasa, al treilea este subsubclasa, al patrulea este numărul de serie al enzimei din această subclasă, de exemplu, codul arginazei este 3.5.3.1.
Mergi la cursurile nr. 2„Structura și funcțiile carbohidraților și lipidelor”
Mergi la cursurile nr. 4„Structura și funcțiile acizilor nucleici ATP”
În prima jumătate a secolului al XIX-lea. mulți chimiști, și printre ei în primul rând J. von Liebig, au ajuns treptat la concluzia că proteinele reprezintă o clasă specială de compuși azotați. Denumirea „proteine” (din greacă.
protos primul) a fost propus în 1840 de chimistul olandez G. Mulder. PROPRIETĂȚI FIZICE Proteinele sunt albe în stare solidă, dar incolore în soluție, cu excepția cazului în care poartă un fel de grup cromofor (colorat), cum ar fi hemoglobina. Solubilitatea în apă variază foarte mult între diferitele proteine. De asemenea, se modifică în funcție de pH și de concentrația sărurilor din soluție, astfel încât este posibil să se selecteze condițiile în care o proteină va precipita selectiv în prezența altor proteine. Această metodă de „sărare” este utilizată pe scară largă pentru izolarea și purificarea proteinelor. Proteina purificată precipită adesea din soluție sub formă de cristale.În comparație cu alți compuși, greutatea moleculară a proteinelor este foarte mare, variind de la câteva mii la multe milioane de daltoni. Prin urmare, în timpul ultracentrifugării, proteinele sunt sedimentate și la rate diferite. Datorită prezenței grupurilor încărcate pozitiv și negativ în moleculele de proteine, acestea se mișcă cu viteze diferite și într-un câmp electric. Aceasta este baza electroforezei, o metodă folosită pentru a izola proteinele individuale din amestecuri complexe. Proteinele sunt de asemenea purificate prin cromatografie.
PROPRIETĂȚI CHIMICE Structura. Proteinele sunt polimeri, adică molecule construite ca lanțuri din unități monomerice repetate sau subunități, al căror rol îl joacă A -aminoacizi. Formula generală a aminoacizilor unde R un atom de hidrogen sau o grupare organică.O moleculă proteică (lanț polipeptidic) poate consta doar dintr-un număr relativ mic de aminoacizi sau câteva mii de unități monomerice. Combinația de aminoacizi într-un lanț este posibilă deoarece fiecare dintre ei are două grupe chimice diferite: o grupă amino cu proprietăți de bază,
NH2 și o grupare carboxil acidă, COOH. Ambele grupuri sunt afiliate A -atomul de carbon. Gruparea carboxil a unui aminoacid poate forma o legătură amidă (peptidă) cu gruparea amino a altui aminoacid: După ce doi aminoacizi au fost legați în acest fel, lanțul poate fi extins prin adăugarea unui al treilea la al doilea aminoacid și așa mai departe. După cum se poate observa din ecuația de mai sus, atunci când se formează o legătură peptidică, este eliberată o moleculă de apă. În prezența acizilor, alcalinelor sau a enzimelor proteolitice, reacția se desfășoară în direcția opusă: lanțul polipeptidic este împărțit în aminoacizi cu adăugarea de apă. Această reacție se numește hidroliză. Hidroliza are loc spontan și este necesară energie pentru a conecta aminoacizii într-un lanț polipeptidic.
După ce doi aminoacizi au fost legați în acest fel, lanțul poate fi extins prin adăugarea unui al treilea la al doilea aminoacid și așa mai departe. După cum se poate observa din ecuația de mai sus, atunci când se formează o legătură peptidică, este eliberată o moleculă de apă. În prezența acizilor, alcalinelor sau a enzimelor proteolitice, reacția se desfășoară în direcția opusă: lanțul polipeptidic este împărțit în aminoacizi cu adăugarea de apă. Această reacție se numește hidroliză. Hidroliza are loc spontan și este necesară energie pentru a conecta aminoacizii într-un lanț polipeptidic. O grupare carboxil și o grupare amidă (sau o grupare imidă similară în cazul aminoacidului prolina) sunt prezente în toți aminoacizii, dar diferențele dintre aminoacizi sunt determinate de natura grupului sau „lanțului lateral”. care este indicat mai sus prin scrisoare
R . Rolul catenei laterale poate fi jucat de un atom de hidrogen, ca în aminoacidul glicină, sau de o grupare voluminoasă, ca în histidină și triptofan. Unele lanțuri laterale sunt inerte din punct de vedere chimic, în timp ce altele sunt semnificativ reactive.Pot fi sintetizați multe mii de aminoacizi diferiți și mulți aminoacizi diferiți apar în natură, dar pentru sinteza proteinelor se folosesc doar 20 de tipuri de aminoacizi: alanină, arginină, asparagină, acid aspartic, valină, histidină, glicină, glutamină, glutamic. acid, izoleucină, leucină, lizină, metionină, prolină, serină, tirozină, treonină, triptofan, fenilalanină și cisteină (în proteine, cisteina poate fi prezentă ca dimer
cistina). Adevărat, unele proteine conțin și alți aminoacizi în plus față de cei douăzeci care apar în mod regulat, dar ele sunt formate ca urmare a modificării unuia dintre cei douăzeci enumerați după ce a fost inclus în proteină.Activitate optică. Toți aminoacizii, cu excepția glicinei, au A Atomul de carbon are patru grupe diferite atașate la el. Din punct de vedere al geometriei, patru grupuri diferite pot fi atașate în două moduri și, în consecință, există două configurații posibile, sau doi izomeri, legați unul de celălalt așa cum este un obiect cu imaginea sa în oglindă, adică. ca mâna stângă spre dreapta. O configurație se numește stânga sau stângaci ( L ), iar celălalt drept, sau dextrogir ( D ), deoarece doi astfel de izomeri diferă în direcția de rotație a planului luminii polarizate. Se găsește doar în proteine L -aminoacizi (excepția este glicina; poate fi reprezentat într-o singură formă, deoarece două dintre cele patru grupe ale sale sunt aceleași), și toți sunt optic activi (din moment ce există un singur izomer). D -aminoacizii sunt rari in natura; se găsesc în unele antibiotice și în peretele celular al bacteriilor.Secvența de aminoacizi. Aminoacizii dintr-un lanț polipeptidic nu sunt aranjați aleatoriu, ci într-o anumită ordine fixă, iar această ordine determină funcțiile și proprietățile proteinei. Variând ordinea celor 20 de tipuri de aminoacizi, puteți crea un număr mare de proteine diferite, la fel cum puteți crea multe texte diferite din literele alfabetului.În trecut, determinarea secvenței de aminoacizi a unei proteine a durat adesea câțiva ani. Determinarea directă este încă o sarcină destul de intensivă în muncă, deși au fost create dispozitive care permit efectuarea acesteia în mod automat. De obicei, este mai ușor să determinați secvența de nucleotide a genei corespunzătoare și să deduceți secvența de aminoacizi a proteinei din aceasta. Până în prezent, secvențele de aminoacizi a multor sute de proteine au fost deja determinate. Funcțiile proteinelor descifrate sunt de obicei cunoscute, iar acest lucru ajută la imaginarea posibilelor funcții ale proteinelor similare formate, de exemplu, în neoplasmele maligne.
Proteine complexe. Proteinele care constau numai din aminoacizi sunt numite simple. Deseori, totuși, un atom de metal sau un compus chimic care nu este un aminoacid este atașat la lanțul polipeptidic. Astfel de proteine sunt numite complexe. Un exemplu este hemoglobina: conține porfirina de fier, care îi determină culoarea roșie și îi permite să acționeze ca purtător de oxigen.Denumirile majorității proteinelor complexe indică natura grupelor atașate: glicoproteinele conțin zaharuri, lipoproteinele conțin grăsimi. Dacă activitatea catalitică a unei enzime depinde de grupul atașat, atunci se numește grup protetic. Adesea, o vitamină joacă rolul unui grup protetic sau face parte din unul. Vitamina A, de exemplu, atașată la una dintre proteinele din retină, determină sensibilitatea acesteia la lumină.
Structura terțiară. Ceea ce este important nu este atât secvența de aminoacizi a proteinei în sine (structura primară), cât și modul în care este așezată în spațiu. De-a lungul întregii lungimi a lanțului polipeptidic, ionii de hidrogen formează legături regulate de hidrogen, care îi conferă forma unui helix sau strat (structură secundară). Din combinarea unor astfel de elice și straturi, reiese o formă compactă de ordinul următor: structura terțiară a proteinei. În jurul legăturilor care țin unitățile monomerice ale lanțului, sunt posibile rotații la unghiuri mici. Prin urmare, din punct de vedere pur geometric, numărul de configurații posibile pentru orice lanț polipeptidic este infinit de mare. În realitate, fiecare proteină există în mod normal într-o singură configurație, determinată de secvența sa de aminoacizi. Această structură nu este rigidă, parcă « respiră” fluctuează în jurul unei anumite configurații medii. Circuitul este pliat într-o configurație în care energia liberă (capacitatea de a produce lucru) este minimă, la fel cum un arc eliberat se comprimă doar într-o stare corespunzătoare energiei libere minime. Adesea, o parte a lanțului este legată rigid de alta prin disulfură ( SS) legături între două reziduuri de cisteină. Acesta este, parțial, motivul pentru care cisteina joacă un rol deosebit de important printre aminoacizi.Complexitatea structurii proteinelor este atât de mare încât nu este încă posibil să se calculeze structura terțiară a unei proteine, chiar dacă secvența ei de aminoacizi este cunoscută. Dar dacă este posibil să se obțină cristale de proteine, atunci structura sa terțiară poate fi determinată prin difracție de raze X.
În proteinele structurale, contractile și în alte proteine, lanțurile sunt alungite și câteva lanțuri ușor pliate aflate în apropiere formează fibrile; fibrilele, la rândul lor, se pliază în formațiuni mai mari de fibre. Cu toate acestea, majoritatea proteinelor în soluție au o formă globulară: lanțurile sunt înfășurate într-un glob, ca firele într-o minge. Energia liberă cu această configurație este minimă, deoarece aminoacizii hidrofobi („resplători de apă”) sunt ascunși în interiorul globului, iar aminoacizii hidrofili („atrage apa”) sunt pe suprafața acestuia.
Multe proteine sunt complexe ale mai multor lanțuri polipeptidice. Această structură se numește structura cuaternară a proteinei. Molecula de hemoglobină, de exemplu, constă din patru subunități, fiecare dintre ele fiind o proteină globulară.
Proteinele structurale, datorită configurației lor liniare, formează fibre care au o rezistență la tracțiune foarte mare, în timp ce configurația globulară permite proteinelor să intre în interacțiuni specifice cu alți compuși. Pe suprafața globului, când lanțurile sunt așezate corect, apar cavități de o anumită formă în care se află grupuri chimice reactive. Dacă o anumită proteină este o enzimă, atunci o altă moleculă, de obicei mai mică, a unei substanțe intră într-o astfel de cavitate, la fel cum o cheie intră într-o lacăt; în acest caz, configurația norului de electroni al moleculei se modifică sub influența grupărilor chimice situate în cavitate, iar acest lucru îl forțează să reacționeze într-un anumit mod. În acest fel, enzima catalizează reacția. Moleculele de anticorpi au, de asemenea, cavități în care se leagă diferite substanțe străine și sunt astfel făcute inofensive. Modelul „lacăt și cheie”, care explică interacțiunea proteinelor cu alți compuși, ne permite să înțelegem specificitatea enzimelor și anticorpilor, adică. capacitatea lor de a reacţiona numai cu anumiţi compuşi.
Proteine în diferite tipuri de organisme. Proteinele care îndeplinesc aceeași funcție la diferite specii de plante și animale și, prin urmare, poartă același nume au, de asemenea, o configurație similară. Ele, totuși, diferă oarecum în secvența lor de aminoacizi. Pe măsură ce speciile se depărtează de un strămoș comun, unii aminoacizi în anumite poziții sunt înlocuiți de mutații cu alții. Mutațiile dăunătoare care provoacă boli ereditare sunt eliminate prin selecție naturală, dar cele benefice sau cel puțin neutre pot persista. Cu cât două specii sunt mai apropiate una de cealaltă, cu atât se găsesc mai puține diferențe în proteinele lor.Unele proteine se schimbă relativ repede, altele sunt foarte conservate. Acesta din urmă include, de exemplu, citocromul Cu o enzimă respiratorie găsită în majoritatea organismelor vii. La oameni și la cimpanzei, secvențele sale de aminoacizi sunt identice și în citocrom Cu La grâu, doar 38% dintre aminoacizi au fost diferiți. Chiar și comparând oameni și bacterii, asemănarea citocromilor Cu(diferențele afectează 65% dintre aminoacizii de aici) încă pot fi văzute, deși strămoșul comun al bacteriilor și al oamenilor a trăit pe Pământ în urmă cu aproximativ două miliarde de ani. În zilele noastre, compararea secvențelor de aminoacizi este adesea folosită pentru a construi un arbore filogenetic (genealogic), care reflectă relațiile evolutive dintre diferite organisme.
Denaturarea. Molecula de proteină sintetizată, pliabilă, capătă configurația sa caracteristică. Această configurație poate fi însă distrusă prin încălzire, prin modificarea pH-ului, prin expunerea la solvenți organici și chiar prin simpla agitare a soluției până când apar bule pe suprafața acesteia. O proteină modificată în acest fel se numește denaturată; își pierde activitatea biologică și devine de obicei insolubilă. Exemple binecunoscute de proteine denaturate sunt ouăle fierte sau frișca. Proteinele mici care conțin doar aproximativ o sută de aminoacizi sunt capabile de renaturare, adică. redobândiți configurația originală. Dar majoritatea proteinelor se transformă pur și simplu într-o masă de lanțuri polipeptidice încurcate și nu își restabilesc configurația anterioară.Una dintre principalele dificultăți în izolarea proteinelor active este sensibilitatea lor extremă la denaturare. Această proprietate a proteinelor își găsește o aplicație utilă în conservarea alimentelor: temperatura ridicată denaturează ireversibil enzimele microorganismelor, iar microorganismele mor.
SINTEZA PROTEINEI Pentru a sintetiza proteine, un organism viu trebuie să aibă un sistem de enzime capabile să unească un aminoacid cu altul. De asemenea, este necesară o sursă de informații pentru a determina ce aminoacizi ar trebui combinați. Întrucât există mii de tipuri de proteine în organism și fiecare dintre ele constă în medie din câteva sute de aminoacizi, informațiile necesare trebuie să fie cu adevărat enorme. Este stocat (similar cu modul în care o înregistrare este stocată pe o bandă magnetică) în moleculele de acid nucleic care alcătuiesc genele. Cm . tot EREDITAR; ACIZI NUCLEICI.Activarea enzimatică. Un lanț polipeptidic sintetizat din aminoacizi nu este întotdeauna o proteină în forma sa finală. Multe enzime sunt sintetizate mai întâi ca precursori inactivi și devin active numai după ce o altă enzimă elimină mai mulți aminoacizi la un capăt al lanțului. Unele dintre enzimele digestive, cum ar fi tripsina, sunt sintetizate în această formă inactivă; aceste enzime sunt activate în tractul digestiv ca urmare a îndepărtării fragmentului terminal al lanțului. Hormonul insulina, a cărei moleculă, în forma sa activă, este formată din două lanțuri scurte, este sintetizat sub forma unui lanț, așa-numitul. proinsulină. Partea de mijloc a acestui lanț este apoi îndepărtată, iar fragmentele rămase se leagă împreună pentru a forma molecula de hormon activ. Proteinele complexe se formează numai după ce o anumită grupă chimică este atașată la proteină, iar această atașare necesită adesea și o enzimă.Circulația metabolică. După hrănirea unui animal cu aminoacizi marcați cu izotopi radioactivi de carbon, azot sau hidrogen, eticheta este rapid încorporată în proteinele sale. Dacă aminoacizii marcați nu mai pătrund în organism, cantitatea de etichetă din proteine începe să scadă. Aceste experimente arată că proteinele rezultate nu sunt reținute în organism până la sfârșitul vieții. Toate, cu puține excepții, sunt într-o stare dinamică, descompunându-se constant în aminoacizi și apoi fiind sintetizate din nou.Unele proteine se descompun atunci când celulele mor și sunt distruse. Acest lucru se întâmplă tot timpul, de exemplu, cu celulele roșii din sânge și celulele epiteliale care căptușesc suprafața interioară a intestinului. În plus, descompunerea și resinteza proteinelor are loc și în celulele vii. Destul de ciudat, se știe mai puțin despre descompunerea proteinelor decât despre sinteza lor. Este clar, totuși, că defalcarea implică enzime proteolitice similare cu cele care descompun proteinele în aminoacizi în tractul digestiv.
Timpul de înjumătățire al diferitelor proteine variază de la câteva ore la multe luni. Singura excepție este molecula de colagen. Odată formate, acestea rămân stabile și nu sunt reînnoite sau înlocuite. În timp, însă, unele dintre proprietățile lor se modifică, în special elasticitatea, și, deoarece nu sunt reînnoite, acest lucru are ca rezultat anumite modificări legate de vârstă, cum ar fi apariția ridurilor pe piele.
Proteine sintetice. Chimiștii au învățat de mult să polimerizeze aminoacizii, dar aminoacizii sunt combinați într-o manieră dezordonată, astfel încât produsele unei astfel de polimerizări se aseamănă puțin cu cele naturale. Adevărat, este posibilă combinarea aminoacizilor într-o ordine dată, ceea ce face posibilă obținerea unor proteine active biologic, în special insulină. Procesul este destul de complicat, iar în acest fel este posibil să se obțină doar acele proteine ale căror molecule conțin aproximativ o sută de aminoacizi. Este de preferat în schimb să se sintetizeze sau să izola secvența de nucleotide a unei gene corespunzătoare secvenței de aminoacizi dorite și apoi să se introducă această genă într-o bacterie, care va produce cantități mari din produsul dorit prin replicare. Această metodă are însă și dezavantajele sale. Cm . de asemenea INGINERIA GENETICĂ. PROTEINE ȘI NUTRIȚIE Când proteinele din organism sunt descompuse în aminoacizi, acești aminoacizi pot fi utilizați din nou pentru a sintetiza proteine. În același timp, aminoacizii înșiși sunt supuși defalcării, deci nu sunt complet reutilizați. De asemenea, este clar că în timpul creșterii, sarcinii și vindecării rănilor, sinteza proteinelor trebuie să depășească descompunerea. Organismul pierde continuu unele proteine; Acestea sunt proteinele părului, ale unghiilor și ale stratului de suprafață al pielii. Prin urmare, pentru a sintetiza proteine, fiecare organism trebuie să primească aminoacizi din alimente. Plantele verzi sintetizează din CO 2 , apa și amoniacul sau nitrații sunt toți cei 20 de aminoacizi care se găsesc în proteine. Multe bacterii sunt, de asemenea, capabile să sintetizeze aminoacizi în prezența zahărului (sau a unui echivalent) și a azotului fixat, dar zahărul este în cele din urmă furnizat de plantele verzi. Animalele au o capacitate limitată de a sintetiza aminoacizi; obțin aminoacizi mâncând plante verzi sau alte animale. În tractul digestiv, proteinele absorbite sunt descompuse în aminoacizi, aceștia din urmă sunt absorbiți și din ele se construiesc proteinele caracteristice unui anumit organism. Niciuna dintre proteinele absorbite nu este încorporată în structurile corpului ca atare. Singura excepție este că la multe mamifere, unii anticorpi materni pot trece intacți prin placentă în fluxul sanguin fetal, iar prin laptele matern (în special la rumegătoare) pot fi transferați nou-născutului imediat după naștere.Necesarul de proteine. Este clar că pentru a menține viața organismul trebuie să primească o anumită cantitate de proteine din alimente. Cu toate acestea, amploarea acestei nevoi depinde de o serie de factori. Organismul are nevoie de hrană atât ca sursă de energie (calorii), cât și ca material pentru construirea structurilor sale. Nevoia de energie este pe primul loc. Aceasta înseamnă că atunci când există puțini carbohidrați și grăsimi în dietă, proteinele dietetice sunt folosite nu pentru sinteza propriilor proteine, ci ca sursă de calorii. În timpul postului prelungit, chiar și propriile proteine sunt folosite pentru a satisface nevoile energetice. Dacă în dietă există destui carbohidrați, atunci consumul de proteine poate fi redus.Bilanțul de azot. În medie cca. 16% din masa totală de proteine este azot. Atunci când aminoacizii conținuti de proteine sunt descompuse, azotul pe care îl conțin este excretat din organism prin urină și (într-o măsură mai mică) în fecale sub formă de diverși compuși azotați. Prin urmare, este convenabil să se utilizeze un indicator precum bilanțul de azot pentru a evalua calitatea nutriției proteice, de exemplu. diferența (în grame) dintre cantitatea de azot care intră în organism și cantitatea de azot excretată pe zi. Cu o alimentație normală la un adult, aceste cantități sunt egale. Într-un organism în creștere, cantitatea de azot excretată este mai mică decât cantitatea primită, adică. soldul este pozitiv. Dacă există o lipsă de proteine în dietă, echilibrul este negativ. Dacă în dietă există suficiente calorii, dar nu există proteine, organismul economisește proteine. În același timp, metabolismul proteic încetinește, iar utilizarea repetată a aminoacizilor în sinteza proteinelor are loc cu cea mai mare eficiență posibilă. Cu toate acestea, pierderile sunt inevitabile, iar compușii azotați sunt încă excretați în urină și parțial în fecale. Cantitatea de azot excretată din organism pe zi în timpul postului proteic poate servi ca măsură a deficienței zilnice de proteine. Este firesc să presupunem că prin introducerea în alimentație a unei cantități de proteine echivalente cu această deficiență, echilibrul de azot poate fi restabilit. Cu toate acestea, nu este. După ce a primit această cantitate de proteine, organismul începe să folosească aminoacizii mai puțin eficient, așa că este nevoie de niște proteine suplimentare pentru a restabili echilibrul de azot.Dacă cantitatea de proteine din dietă depășește ceea ce este necesar pentru a menține echilibrul de azot, atunci se pare că nu există niciun rău. Aminoacizii în exces sunt pur și simplu folosiți ca sursă de energie. Ca un exemplu deosebit de izbitor, eschimosii consumă puțini carbohidrați și de aproximativ zece ori cantitatea de proteine necesară pentru a menține echilibrul de azot. În majoritatea cazurilor, totuși, utilizarea proteinelor ca sursă de energie nu este benefică, deoarece o anumită cantitate de carbohidrați poate produce mult mai multe calorii decât aceeași cantitate de proteine. În țările sărace, oamenii își iau caloriile din carbohidrați și consumă cantități minime de proteine.
Dacă organismul primește numărul necesar de calorii sub formă de produse neproteice, atunci cantitatea minimă de proteine pentru a asigura menținerea echilibrului de azot este de cca. 30 g pe zi. Cam atât de multă proteină este conținută în patru felii de pâine sau 0,5 litri de lapte. Un număr puțin mai mare este de obicei considerat optim; Se recomandă 50 până la 70 g.
Aminoacizi esentiali. Până acum, proteinele erau considerate ca un întreg. Între timp, pentru ca sinteza proteinelor să aibă loc, toți aminoacizii necesari trebuie să fie prezenți în organism. Corpul animalului însuși este capabil să sintetizeze unii dintre aminoacizi. Se numesc înlocuibile deoarece nu trebuie neapărat să fie prezente în dietă, este important doar ca aportul total de proteine ca sursă de azot să fie suficient; apoi, dacă există un deficit de aminoacizi neesențiali, organismul îi poate sintetiza în detrimentul celor care sunt prezenți în exces. Restul de aminoacizi „esențiali” nu pot fi sintetizați și trebuie să fie furnizați organismului prin alimente. Esențiale pentru oameni sunt valina, leucina, izoleucina, treonina, metionina, fenilalanina, triptofanul, histidina, lizina și arginina. (Deși arginina poate fi sintetizată în organism, este clasificată ca un aminoacid esențial deoarece nu este produsă în cantități suficiente la nou-născuți și copiii în creștere. Pe de altă parte, unii dintre acești aminoacizi din alimente pot deveni inutili pentru un adult. persoană.)Această listă de aminoacizi esențiali este aproximativ aceeași la alte vertebrate și chiar la insecte. Valoarea nutritivă a proteinelor este de obicei determinată prin hrănirea lor la șobolani în creștere și monitorizarea creșterii în greutate a animalelor.
Valoarea nutritivă a proteinelor. Valoarea nutritivă a unei proteine este determinată de aminoacidul esențial care este cel mai deficitar. Să ilustrăm acest lucru cu un exemplu. Proteinele din corpul nostru conțin în medie cca. 2% triptofan (în greutate). Să presupunem că dieta include 10 g de proteine care conțin 1% triptofan și că există destui alți aminoacizi esențiali în ea. În cazul nostru, 10 g din această proteină incompletă este în esență echivalent cu 5 g de proteină completă; restul de 5 g poate servi doar ca sursă de energie. Rețineți că, deoarece aminoacizii practic nu sunt stocați în organism și, pentru ca sinteza proteinelor să aibă loc, toți aminoacizii trebuie să fie prezenți în același timp, efectul aportului de aminoacizi esențiali poate fi detectat doar dacă toți intră în corp în același timp. Compoziția medie a majorității proteinelor animale este apropiată de compoziția medie a proteinelor din corpul uman, așa că este puțin probabil să ne confruntăm cu deficiența de aminoacizi dacă dieta noastră este bogată în alimente precum carne, ouă, lapte și brânză. Cu toate acestea, există proteine, precum gelatina (un produs al denaturarii colagenului), care conțin foarte puțini aminoacizi esențiali. Proteinele vegetale, deși sunt mai bune decât gelatina în acest sens, sunt și ele sărace în aminoacizi esențiali; Sunt deosebit de săraci în lizină și triptofan. Cu toate acestea, o dietă pur vegetariană nu poate fi considerată deloc dăunătoare, decât dacă consumă o cantitate ceva mai mare de proteine vegetale, suficientă pentru a furniza organismului aminoacizi esențiali. Plantele conțin cele mai multe proteine în semințele lor, în special în semințele de grâu și diferite leguminoase. Lăstarii tineri, cum ar fi sparanghelul, sunt, de asemenea, bogați în proteine.Proteine sintetice în dietă. Prin adăugarea unor cantități mici de aminoacizi esențiali sintetici sau proteine bogate în aminoacizi la proteinele incomplete, cum ar fi proteinele din porumb, valoarea nutrițională a acestora din urmă poate fi crescută semnificativ, de exemplu. crescând astfel cantitatea de proteine consumată. O altă posibilitate este să crească bacterii sau drojdii pe hidrocarburi petroliere cu adăugarea de nitrați sau amoniac ca sursă de azot. Proteina microbiană obținută în acest mod poate servi ca hrană pentru păsări de curte sau animale, sau poate fi consumată direct de oameni. A treia metodă, utilizată pe scară largă, utilizează fiziologia rumegătoarelor. La rumegătoare, în partea inițială a stomacului, așa-numitele. Rumenul este locuit de forme speciale de bacterii și protozoare care transformă proteinele vegetale incomplete în proteine microbiene mai complete, iar acestea, la rândul lor, după digestie și absorbție se transformă în proteine animale. Ureea, un compus sintetic ieftin care conține azot, poate fi adăugată în hrana animalelor. Microorganismele care trăiesc în rumen folosesc azotul ureic pentru a transforma carbohidrații (din care sunt mult mai mulți în furaj) în proteine. Aproximativ o treime din tot azotul din hrana animalelor poate veni sub formă de uree, ceea ce înseamnă în esență, într-o anumită măsură, sinteza chimică a proteinelor. În SUA, această metodă joacă un rol important ca una dintre modalitățile de obținere a proteinelor.LITERATURĂ Murray R., Grenner D., Mayes P., Rodwell W. Biochimia umană, vol. 12. M., 1993Alberts B, Bray D, Lewis J, și colab. Biologie celulară moleculară, vol. 13. M., 1994
Biletul 2. 1. Factori nutritivi esentiali de natura lipidica. Unele lipide nu sunt sintetizate în corpul uman și, prin urmare, sunt factori nutriționali esențiali. Acestea includ acizi grași cu două sau mai multe legături duble (polien) - acizi grași esențiali. Unii dintre acești acizi sunt substraturi pentru sinteza hormonilor locali - eicosanoizi (subiectul 8.10).
Vitamine liposolubile indeplineste diverse functii: vitamina A participă la procesul de vedere, precum și la creșterea și diferențierea celulelor; capacitatea sa de a inhiba creșterea anumitor tipuri de tumori a fost dovedită; vitamina K participă la coagularea sângelui; vitamina D participă la reglarea metabolismului calciului; vitamina E- antioxidant, inhiba formarea radicalilor liberi si astfel contracareaza deteriorarea celulelor ca urmare a peroxidarii lipidelor.
Document
2.Structura și nivelurile de organizare structurală a proteinelor
Există patru niveluri de organizare structurală a proteinelor: primar, secundar, terțiar și cuaternar. Fiecare nivel are propriile sale caracteristici.
Structura primară a proteinei
Structura primară a proteinelor este un lanț polipeptidic liniar de aminoacizi conectați prin legături peptidice. Structura primară este cel mai simplu nivel de organizare structurală a unei molecule de proteine. Stabilitate ridicată îi este dată de legăturile peptidice covalente dintre gruparea α-amino a unui aminoacid și gruparea α-carboxil a unui alt aminoacid. [spectacol].
Dacă gruparea imino a prolinei sau hidroxiprolinei este implicată în formarea unei legături peptidice, atunci are o formă diferită [spectacol].
Când se formează legături peptidice în celule, gruparea carboxil a unui aminoacid este mai întâi activată și apoi se combină cu gruparea amino a altuia. Sinteza de laborator a polipeptidelor se realizează aproximativ în același mod.
O legătură peptidică este un fragment repetat al unui lanț polipeptidic. Are o serie de caracteristici care afectează nu numai forma structurii primare, ci și nivelurile superioare de organizare a lanțului polipeptidic:

coplanaritate - toți atomii incluși în grupul peptidic sunt în același plan;
capacitatea de a exista în două forme de rezonanță (forma ceto sau enol);
poziţia trans a substituenţilor faţă de legătura C-N;
capacitatea de a forma legături de hidrogen și fiecare dintre grupele peptidice poate forma două legături de hidrogen cu alte grupări, inclusiv cu cele peptidice.
Excepție fac grupele peptidice care implică gruparea amino a prolinei sau hidroxiprolinei. Ei sunt capabili să formeze o singură legătură de hidrogen (vezi mai sus). Acest lucru afectează formarea structurii secundare a proteinei. Lanțul polipeptidic din zona în care se află prolina sau hidroxiprolina se îndoaie ușor, deoarece nu este ținut, ca de obicei, de o a doua legătură de hidrogen.
Nomenclatura peptidelor și polipeptidelor. Numele peptidelor este alcătuit din denumirile aminoacizilor lor constituenți. Doi aminoacizi formează o dipeptidă, trei fac o tripeptidă, patru fac o tetrapeptidă etc. Fiecare peptidă sau lanț polipeptidic de orice lungime are un aminoacid N-terminal care conține o grupare amino liberă și un aminoacid C-terminal care conține un carboxil liber. grup. La denumirea polipeptidelor, toţi aminoacizii sunt enumeraţi secvenţial, începând cu cel N-terminal, înlocuind în denumirea lor, cu excepţia celui C-terminal, sufixul -in cu -il (întrucât aminoacizii din peptide nu mai au un grupa carboxil, ci una carbonil). De exemplu, numele prezentat în Fig. 1 tripeptidă - leuc nămol fenilalan nămol treon în.
Caracteristicile structurii primare a proteinei. În coloana vertebrală a lanțului polipeptidic, structurile rigide (grupe peptidice plate) alternează cu regiuni relativ mobile (-CHR), care sunt capabile să se rotească în jurul legăturilor. Astfel de caracteristici structurale ale lanțului polipeptidic afectează aranjamentul său spațial.
Structura secundară a proteinei
Structura secundară este o modalitate de pliere a unui lanț polipeptidic într-o structură ordonată datorită formării legăturilor de hidrogen între grupările peptidice ale aceluiași lanț sau lanțurile polipeptidice adiacente. În funcție de configurația lor, structurile secundare sunt împărțite în elicoidale (α-helix) și pliate în straturi (β-structură și cruce-β-form).


α-helix. Acesta este un tip de structură proteică secundară care arată ca o spirală obișnuită, formată datorită legăturilor de hidrogen interpeptidice din cadrul unui lanț polipeptidic. Modelul structurii α-helixului (Fig. 2), care ia în considerare toate proprietățile legăturii peptidice, a fost propus de Pauling și Corey. Principalele caracteristici ale α-helix:
configurația elicoidală a lanțului polipeptidic având simetrie elicoidală;
formarea de legături de hidrogen între grupările peptidice ale fiecărui prim și al patrulea rest de aminoacid;
regularitatea rotațiilor spiralate;
echivalența tuturor reziduurilor de aminoacizi din α-helix, indiferent de structura radicalilor lor laterali;
radicalii laterali ai aminoacizilor nu participă la formarea α-helixului.
În exterior, α-helixul arată ca o spirală ușor întinsă a unui aragaz electric. Regularitatea legăturilor de hidrogen între prima și a patra grupare peptidică determină regularitatea spirelor lanțului polipeptidic. Înălțimea unei ture sau pasul elicei α este de 0,54 nm; include 3,6 resturi de aminoacizi, adică fiecare reziduu de aminoacizi se mișcă de-a lungul axei (înălțimea unui rest de aminoacizi) cu 0,15 nm (0,54:3,6 = 0,15 nm), ceea ce ne permite să vorbim despre echivalența tuturor reziduurilor de aminoacizi. în α-helix. Perioada de regularitate a unui α-helix este de 5 ture sau 18 reziduuri de aminoacizi; lungimea unei perioade este de 2,7 nm. Orez. 3. Modelul a-helix Pauling-Corey
β-Structură. Acesta este un tip de structură secundară care are o configurație ușor curbată a lanțului polipeptidic și este format din legături interpeptidice de hidrogen în secțiuni individuale ale unui lanț polipeptidic sau lanțuri polipeptidice adiacente. Se mai numește și o structură cu pliuri stratificate. Există varietăți de structuri β. Regiunile limitate stratificate formate dintr-un lanț polipeptidic al unei proteine sunt numite formă încrucișată β (structură β scurtă). Legăturile de hidrogen în formă încrucișată-β se formează între grupările peptidice ale buclelor lanțului polipeptidic. Un alt tip - structura β completă - este caracteristic întregului lanț polipeptidic, care are o formă alungită și este ținut de legături interpeptidice de hidrogen între lanțurile polipeptidice paralele adiacente (Fig. 3). Această structură seamănă cu burduful unui acordeon. Mai mult, sunt posibile variante ale structurilor β: pot fi formate din lanțuri paralele (capetele N-terminale ale lanțurilor polipeptidice sunt direcționate în aceeași direcție) și antiparalele (capetele N-terminale sunt direcționate în direcții diferite). Radicalii laterali ai unui strat sunt plasați între radicalii laterali ai altui strat.
În proteine, tranzițiile de la structurile α la structurile β și înapoi sunt posibile datorită rearanjarii legăturilor de hidrogen. În loc de legături interpeptidice regulate de hidrogen de-a lungul lanțului (mulțumită cărora lanțul polipeptidic este răsucit într-o spirală), secțiunile elicoidale se desfășoară și legăturile de hidrogen se închid între fragmentele alungite ale lanțurilor polipeptidice. Această tranziție se găsește în keratina, proteina părului. La spălarea părului cu detergenți alcalini, structura elicoidală a β-keratinei este ușor distrusă și se transformă în α-keratina (părul creț se îndreaptă).
Distrugerea structurilor secundare regulate ale proteinelor (elice α și structuri β), prin analogie cu topirea unui cristal, se numește „topirea” polipeptidelor. În acest caz, legăturile de hidrogen sunt rupte, iar lanțurile polipeptidice iau forma unei încurcături aleatorii. În consecință, stabilitatea structurilor secundare este determinată de legăturile de hidrogen interpeptidice. Alte tipuri de legături nu participă aproape deloc la aceasta, cu excepția legăturilor disulfurice de-a lungul lanțului polipeptidic în locațiile reziduurilor de cisteină. Peptidele scurte sunt închise în cicluri din cauza legăturilor disulfurice. Multe proteine conțin atât regiuni elicoidale α, cât și structuri β. Aproape nu există proteine naturale formate din 100% α-helix (excepția este paramiozina, o proteină musculară care este 96-100% α-helix), în timp ce polipeptidele sintetice au 100% helix.
Alte proteine au grade diferite de înfăşurare. O frecvență ridicată a structurilor elicoidale α este observată în paramiozină, mioglobină și hemoglobină. În contrast, în tripsină, o ribonuclează, o parte semnificativă a lanțului polipeptidic este pliată în structuri β stratificate. Proteinele țesuturilor de susținere: keratina (proteina părului, lână), colagenul (proteina tendoanelor, pielea), fibroina (proteina din mătasea naturală) au o configurație β a lanțurilor polipeptidice. Gradele diferite de elicitate ale lanțurilor polipeptidice ale proteinelor indică faptul că, în mod evident, există forțe care perturbă parțial helicitatea sau „rup” plierea regulată a lanțului polipeptidic. Motivul pentru aceasta este o pliere mai compactă a lanțului polipeptidic proteic într-un anumit volum, adică într-o structură terțiară.
Structura terțiară a proteinelor
Structura terțiară a unei proteine este modul în care lanțul polipeptidic este aranjat în spațiu. Pe baza formei structurii lor terțiare, proteinele sunt împărțite în principal în globulare și fibrilare. Proteinele globulare au cel mai adesea o formă elipsoidă, iar proteinele fibrilare (sub formă de fir) au o formă alungită (forma tijă sau fus).
Cu toate acestea, configurația structurii terțiare a proteinelor nu dă încă motive să se creadă că proteinele fibrilare au doar o structură β, iar proteinele globulare au o structură α-helidiană. Există proteine fibrilare care au o structură secundară pliată, mai degrabă elicoidală decât stratificată. De exemplu, α-keratina și paramiozina (proteina mușchiului obturator al moluștelor), tropomiozinele (proteinele mușchilor scheletici) aparțin proteinelor fibrilare (au formă de tijă), iar structura lor secundară este α-helix; în contrast, proteinele globulare pot conține un număr mare de structuri β.
Spiralizarea unui lanț polipeptidic linear reduce dimensiunea acestuia de aproximativ 4 ori; iar ambalarea în structura terțiară o face de zeci de ori mai compactă decât lanțul original.
Legături care stabilizează structura terțiară a unei proteine. Legăturile dintre radicalii laterali ai aminoacizilor joacă un rol în stabilizarea structurii terțiare. Aceste conexiuni pot fi împărțite în:
puternic (covalent) [spectacol].
slab (polar și van der Waals) [spectacol].
Numeroase legături între radicalii laterali ai aminoacizilor determină configurația spațială a moleculei proteice.
Caracteristicile organizării structurii terțiare a proteinelor. Conformația structurii terțiare a lanțului polipeptidic este determinată de proprietățile radicalilor laterali ai aminoacizilor incluși în acesta (care nu au un efect vizibil asupra formării structurilor primare și secundare) și de micromediu, adică mediu inconjurator. Atunci când este pliat, lanțul polipeptidic al unei proteine tinde să ia o formă favorabilă din punct de vedere energetic, caracterizată printr-un minim de energie liberă. Prin urmare, grupările R nepolare, „evitând” apa, formează, așa cum ar fi, partea internă a structurii terțiare a proteinei, unde se află partea principală a reziduurilor hidrofobe ale lanțului polipeptidic. Aproape că nu există molecule de apă în centrul globului proteic. Grupările R polare (hidrofile) ale aminoacidului sunt situate în afara acestui miez hidrofob și sunt înconjurate de molecule de apă. Lanțul polipeptidic este îndoit complex în spațiul tridimensional. Când se îndoaie, conformația elicoială secundară este perturbată. Lanțul „se rupe” în punctele slabe în care se află prolina sau hidroxiprolina, deoarece acești aminoacizi sunt mai mobili în lanț, formând o singură legătură de hidrogen cu alte grupări peptidice. Un alt loc de îndoire este glicina, care are o grupă R mică (hidrogen). Prin urmare, grupele R ale altor aminoacizi, atunci când sunt stivuite, tind să ocupe spațiul liber din locația glicinei. O serie de aminoacizi - alanina, leucina, glutamatul, histidina - contribuie la pastrarea structurilor elicoidale stabile in proteine, iar cum ar fi metionina, valina, izoleucina, acidul aspartic favorizeaza formarea structurilor β. Într-o moleculă de proteină cu o configurație terțiară, există regiuni sub formă de elice α (elicoidale), structuri β (stratificate) și o spirală aleatorie. Numai aranjarea spațială corectă a proteinei o face activă; încălcarea acestuia duce la modificări ale proprietăților proteinelor și la pierderea activității biologice.
Structura proteinelor cuaternare
Proteinele formate dintr-un lanț polipeptidic au doar structură terțiară. Acestea includ mioglobina - o proteină din țesutul muscular implicată în legarea oxigenului, o serie de enzime (lizozimă, pepsină, tripsină etc.). Cu toate acestea, unele proteine sunt construite din mai multe lanțuri polipeptidice, fiecare dintre ele având o structură terțiară. Pentru astfel de proteine a fost introdus conceptul de structură cuaternară, care este organizarea mai multor lanțuri polipeptidice cu structură terțiară într-o singură moleculă de proteină funcțională. O astfel de proteină cu structură cuaternară se numește oligomer, iar lanțurile sale polipeptidice cu structură terțiară se numesc protomeri sau subunități (Fig. 4).

La nivel cuaternar de organizare, proteinele păstrează configurația de bază a structurii terțiare (globulare sau fibrilare). De exemplu, hemoglobina este o proteină cu structură cuaternară și este formată din patru subunități. Fiecare dintre subunități este o proteină globulară și, în general, hemoglobina are și o configurație globulară. Proteinele părului și lânii - keratinele, înrudite în structură terțiară cu proteinele fibrilare, au o conformație fibrilă și o structură cuaternară.
Stabilizarea structurii cuaternare a proteinelor. Toate proteinele care au o structură cuaternară sunt izolate sub formă de macromolecule individuale care nu se descompun în subunități. Contactele dintre suprafețele subunităților sunt posibile numai datorită grupărilor polare ale reziduurilor de aminoacizi, deoarece în timpul formării structurii terțiare a fiecăruia dintre lanțurile polipeptidice, radicalii laterali ai aminoacizilor nepolari (care alcătuiesc majoritatea toți aminoacizii proteinogeni) sunt ascunși în interiorul subunității. Între grupările lor polare se formează numeroase legături ionice (sare), hidrogen și, în unele cazuri, legături disulfurice, care țin ferm subunitățile sub forma unui complex organizat. Utilizarea de substanțe care rup legăturile de hidrogen sau de substanțe care reduc punțile disulfurice determină dezagregarea protomerilor și distrugerea structurii cuaternare a proteinei. În tabel 1 rezumă datele privind legăturile care stabilizează diferite niveluri de organizare a moleculei proteice [spectacol].
Caracteristici ale organizării structurale a unor proteine fibrilare
Organizarea structurală a proteinelor fibrilare are o serie de caracteristici în comparație cu proteinele globulare. Aceste caracteristici pot fi observate în exemplul keratinei, fibroinei și colagenului. Keratinele există în formațiile α și β. α-Keratinele și fibroina au o structură secundară stratificată, totuși, în cheratina lanțurile sunt paralele, iar în fibroină sunt antiparalele (vezi Fig. 3); În plus, keratina conține legături disulfurice intercatenare, în timp ce fibroina nu le are. Ruperea legăturilor disulfurice duce la separarea lanțurilor polipeptidice din cheratine. Dimpotrivă, formarea numărului maxim de legături disulfurice în cheratine prin expunerea la agenți oxidanți creează o structură spațială puternică. În general, în proteinele fibrilare, spre deosebire de proteinele globulare, uneori este dificil să se facă distincția strictă între diferitele niveluri de organizare. Dacă acceptăm (ca pentru o proteină globulară) că structura terțiară ar trebui să fie formată prin așezarea unui lanț polipeptidic în spațiu, iar structura cuaternară cu mai multe lanțuri, atunci în proteinele fibrilare sunt implicate mai multe lanțuri polipeptidice deja în timpul formării structurii secundare. . Un exemplu tipic de proteină fibrilă este colagenul, care este una dintre cele mai abundente proteine din corpul uman (aproximativ 1/3 din masa tuturor proteinelor). Se găsește în țesuturile care au rezistență mare și extensibilitate scăzută (oase, tendoane, piele, dinți etc.). În colagen, o treime din reziduurile de aminoacizi sunt glicină și aproximativ un sfert sau puțin mai mult sunt prolină sau hidroxiprolină.

Lanțul polipeptidic izolat de colagen (structură primară) arată ca o linie întreruptă. Conține aproximativ 1000 de aminoacizi și are o greutate moleculară de aproximativ 10 5 (Fig. 5, a, b). Lanțul polipeptidic este construit dintr-un trio repetat de aminoacizi (triplet) cu următoarea compoziție: gly-A-B, unde A și B sunt orice aminoacizi, alții decât glicina (cel mai adesea prolină și hidroxiprolina). Lanțurile polipeptidice de colagen (sau lanțuri α) în timpul formării structurilor secundare și terțiare (Fig. 5, c și d) nu pot produce elice α tipice cu simetrie elicoidală. Prolina, hidroxiprolina și glicina (aminoacizi antihelical) interferează cu acest lucru. Prin urmare, trei lanțuri α formează, parcă, spirale răsucite, ca trei fire care se înfășoară în jurul unui cilindru. Trei lanțuri α elicoidale formează o structură de colagen care se repetă numită tropocolagen (Fig. 5d). Tropocolagenul în organizarea sa este structura terțiară a colagenului. Inelele plate de prolină și hidroxiprolină care alternează în mod regulat de-a lungul lanțului îi conferă rigiditate, la fel ca și legăturile intercatenare dintre lanțurile α de tropocolagen (de aceea colagenul este rezistent la întindere). Tropocolagenul este în esență o subunitate a fibrilelor de colagen. Așezarea subunităților de tropocolagen în structura cuaternară a colagenului are loc în mod treptat (Fig. 5e).
Stabilizarea structurilor de colagen are loc datorită legăturilor intercatenare de hidrogen, ionice și van der Waals și unui număr mic de legături covalente.
Lanțurile α de colagen au structuri chimice diferite. Există diferite tipuri de lanțuri α 1 (I, II, III, IV) și lanțuri α 2. În funcție de care lanțuri α 1 - și α 2 - sunt implicate în formarea helixului cu trei catene de tropocolagen, se disting patru tipuri de colagen:
primul tip - două α 1 (I) și un lanț α 2;
al doilea tip - trei lanțuri α 1 (II);
al treilea tip - trei lanțuri α 1 (III);
al patrulea tip - trei lanțuri α 1 (IV).
Cel mai frecvent colagen este primul tip: se găsește în țesutul osos, piele, tendoane; colagenul de tip 2 se găsește în țesutul cartilajului etc. Un tip de țesut poate conține diferite tipuri de colagen.
Agregarea ordonată a structurilor de colagen, rigiditatea și inerția lor asigură rezistența ridicată a fibrelor de colagen. Proteinele de colagen conțin și componente carbohidrați, adică sunt complexe proteine-carbohidrați.
Colagenul este o proteină extracelulară care este formată din celulele țesutului conjunctiv găsite în toate organele. Prin urmare, cu deteriorarea colagenului (sau întreruperea formării acestuia), apar multiple încălcări ale funcțiilor de susținere ale țesutului conjunctiv al organelor.
Lanțul polipeptidic alfa se termină cu combinația de aminoacizi valină-leucină, iar lanțul polipeptidic beta se termină cu combinația valină-histidină-leucină. Lanțurile polipeptidice alfa și beta din molecula de hemoglobină nu sunt aranjate liniar; aceasta este structura primară. Datorită existenței forțelor intramoleculare, lanțurile polipeptidice sunt răsucite sub forma unei helix alfa-helix (structură secundară) tipică proteinelor. Helixul alfa-helix în sine se îndoaie spațial pentru fiecare lanț polipeptidic alfa și beta, formând plexuri de formă ovoidă (structură terțiară). Părțile individuale ale spiralelor alfa-helix ale lanțurilor polipeptidice sunt marcate cu litere latine de la A la H. Toate cele patru lanțuri polipeptidice alfa și beta curbate terțiare sunt situate spațial într-o anumită relație - o structură cuaternară. Ele sunt conectate nu prin legături chimice reale, ci prin forțe intermoleculare.
S-a dovedit că oamenii au trei tipuri principale de hemoglobină normală: embrionară - U, fetală - F și hemoglobină adultă - A. HbU (numit după litera inițială a cuvântului uter) apare în embrion între 7 și 12 săptămâni de viață, apoi dispare si apare hemoglobina fetala, care dupa luna a treia este hemoglobina principala a fatului. În continuare, apare treptat hemoglobina adultă obișnuită, numită HbA, după litera inițială a cuvântului englez „adult”. Cantitatea de hemoglobină fetală scade treptat, astfel încât în momentul nașterii, 80% din hemoglobină este HbA și doar 20% este HbF. După naștere, hemoglobina fetală continuă să scadă și la 2–3 ani de viață este de doar 1–2%. Aceeași cantitate de hemoglobină fetală este la un adult. O cantitate de HbF care depășește 2% este considerată patologică pentru un adult și pentru copiii cu vârsta peste 3 ani.
Pe lângă tipurile normale de hemoglobină, în prezent sunt cunoscute peste 50 de variante patologice. Au fost numiți pentru prima dată cu litere latine. Litera B este absentă în desemnarea tipurilor de hemoglobină, deoarece inițial a desemnat HbS.
Hemoglobina (Hb)- o cromoproteină prezentă în celulele roșii din sânge și implicată în transportul oxigenului către țesuturi. Hemoglobina la adulți se numește hemoglobină A (Hb A). Greutatea sa moleculară este de aproximativ 65.000 Da. Molecula Hb A are o structură cuaternară și include patru subunități - lanțuri polipeptidice (denumite α1, α2, β1 și β2, fiecare dintre ele asociată cu hem.
Amintiți-vă că hemoglobina este o proteină alosterică; moleculele sale se pot schimba reversibil de la o conformație la alta. Aceasta modifică afinitatea proteinei pentru liganzi. Conformația cu cea mai mică afinitate pentru ligand se numește timp sau T-conformație. Conformația cu cea mai mare afinitate pentru ligand este denumită conformație relaxată sau R-conformație.
Conformațiile R și T ale moleculei de hemoglobină sunt într-o stare de echilibru dinamic:
Diferiți factori de mediu pot schimba acest echilibru într-o direcție sau alta. Regulatorii alosterici care afectează afinitatea Hb pentru O2 sunt: 1) oxigenul; 2) concentraţia H+ (pH mediu); 3) dioxid de carbon (CO2); 4) 2,3-difosfoglicerat (DPG). Atașarea unei molecule de oxigen la una dintre subunitățile hemoglobinei promovează tranziția unei conformații tensionate la una relaxată și crește afinitatea pentru oxigen a altor subunități ale aceleiași molecule de hemoglobină. Acest fenomen se numește efect de cooperare. Natura complexă a legării hemoglobinei de oxigen este reflectată de curba de saturație a hemoglobinei O2, care are o formă de S (Figura 3.1).

Figura 3.1. Curbele de saturație a mioglobinei (1) și hemoglobinei (2).
Forme moleculare hemoglobina diferă între ele în structura lanțurilor polipeptidice. Un exemplu de astfel de varietate de hemoglobină care există în condiții fiziologice este hemoglobina fetală (HbF), prezente în sânge în timpul etapei embrionare a dezvoltării umane. Spre deosebire de HbA, molecula sa conține 2 lanțuri α și 2 lanțuri γ (adică lanțurile β sunt înlocuite cu lanțuri γ). O astfel de hemoglobină are o afinitate mai mare pentru oxigen. Acesta este ceea ce permite embrionului să primească oxigen din sângele mamei prin placentă. La scurt timp după naștere, HbF din sângele copilului este înlocuită cu HbA.
Un exemplu de hemoglobină anormală sau patologică este hemoglobina S deja menționată (vezi 2.4.), întâlnită la pacienții cu anemie falciformă. După cum știți deja, aceasta diferă de hemoglobina A prin înlocuirea glutamatului cu valină în lanțurile β. Această substituție de aminoacizi determină o scădere a solubilității HbS în apă și o scădere a afinității sale pentru O2.
După cum știți, proteinele sunt baza pentru originea vieții pe planeta noastră. Dar picătura de coacervat, constând din molecule peptidice, a devenit baza pentru originea viețuitoarelor. Acest lucru este fără îndoială, deoarece analiza compoziției interne a oricărui reprezentant al biomasei arată că aceste substanțe sunt prezente în orice: plante, animale, microorganisme, ciuperci, viruși. În plus, sunt foarte diverse și de natură macromoleculară.
Aceste structuri au patru nume, toate fiind sinonime:
- proteine;
- proteine;
- polipeptide;
- peptide.
Molecule de proteine
Numărul lor este cu adevărat nenumărat. În acest caz, toate moleculele de proteine pot fi împărțite în două grupuri mari:
- simplu - constau numai din secvențe de aminoacizi legate prin legături peptidice;
- complex - structura și structura proteinei sunt caracterizate de grupuri protolitice (protetice) suplimentare, numite și cofactori.
În același timp, moleculele complexe au și propria lor clasificare.
Gradarea peptidelor complexe
- Glicoproteinele sunt compuși strâns legați de proteine și carbohidrați. Grupele protetice de mucopolizaharide sunt țesute în structura moleculei.
- Lipoproteinele sunt un compus complex de proteine și lipide.
- Metaloproteinele - ionii metalici (fier, mangan, cupru și altele) acționează ca un grup protetic.
- Nucleoproteinele sunt legătura dintre proteine și acizi nucleici (ADN, ARN).
- Fosfoproteine - conformația unei proteine și a unui rest de acid ortofosforic.
- Cromoproteinele sunt foarte asemănătoare cu metaloproteinele, totuși, elementul care face parte din grupul protetic este un complex colorat întreg (roșu - hemoglobină, verde - clorofilă și așa mai departe).
În fiecare grup luat în considerare, structura și proprietățile proteinelor sunt diferite. Funcțiile pe care le îndeplinesc variază și în funcție de tipul de moleculă.

Structura chimică a proteinelor
Din acest punct de vedere, proteinele sunt un lanț lung și masiv de reziduuri de aminoacizi conectate între ele prin legături specifice numite legături peptidice. Ramuri numite radicali se extind din structurile laterale ale acizilor. Această structură moleculară a fost descoperită de E. Fischer la începutul secolului XXI.
Ulterior, proteinele, structura și funcțiile proteinelor au fost studiate mai detaliat. A devenit clar că există doar 20 de aminoacizi care formează structura peptidei, dar aceștia pot fi combinați într-o varietate de moduri. De aici varietatea structurilor polipeptidice. În plus, în procesul vieții și în îndeplinirea funcțiilor lor, proteinele sunt capabile să sufere o serie de transformări chimice. Drept urmare, acestea schimbă structura și apare un tip complet nou de conexiune.
Pentru a rupe legătura peptidică, adică pentru a perturba proteina și structura lanțurilor, trebuie să selectați condiții foarte stricte (temperaturi ridicate, acizi sau alcalii, un catalizator). Acest lucru se datorează rezistenței ridicate a moleculei, și anume în grupul peptidic.

Detectarea structurii proteinei în laborator se realizează folosind reacția biuret - expunere la polipeptidă proaspăt precipitată (II). Complexul grupului de peptide și ionul de cupru dă o culoare violet strălucitor.
Există patru organizații structurale principale, fiecare dintre ele având propriile caracteristici structurale ale proteinelor.
Niveluri de organizare: structura primară
După cum sa menționat mai sus, o peptidă este o secvență de resturi de aminoacizi cu sau fără incluziuni, coenzime. Deci, principalul este structura unei molecule care sunt naturali, naturali, cu adevărat aminoacizi conectați prin legături peptidice și nimic mai mult. Adică o polipeptidă cu structură liniară. Mai mult, caracteristicile structurale ale proteinelor de acest tip sunt că o astfel de combinație de acizi este decisivă pentru îndeplinirea funcțiilor moleculei proteice. Datorită prezenței acestor caracteristici, este posibil nu numai să se identifice o peptidă, ci și să se prezică proprietățile și rolul uneia complet noi, încă nedescoperite. Exemple de peptide cu o structură primară naturală sunt insulina, pepsina, chimotripsina și altele.

Conformatia secundara
Structura și proprietățile proteinelor din această categorie variază oarecum. O astfel de structură poate fi formată inițial prin natură sau atunci când cea primară este expusă la hidroliză severă, temperatură sau alte condiții.
Această conformație are trei soiuri:
- Viraje netede, regulate, stereoregulate, construite din reziduuri de aminoacizi, care se răsucesc în jurul axei principale a conexiunii. Ele sunt ținute împreună numai de cele care apar între oxigenul unei grupe peptidice și hidrogenul alteia. Mai mult, structura este considerată corectă datorită faptului că turele se repetă uniform la fiecare 4 verigi. O astfel de structură poate fi fie stângaci, fie dreptaci. Dar în majoritatea proteinelor cunoscute predomină izomerul dextrogiro. Astfel de conformații sunt de obicei numite structuri alfa.
- Compoziția și structura proteinelor de tipul următor diferă de cea precedentă prin aceea că legăturile de hidrogen nu se formează între reziduurile adiacente unei părți a moleculei, ci între cele semnificativ îndepărtate și la o distanță destul de mare. Din acest motiv, întreaga structură ia forma mai multor lanțuri polipeptidice ondulate, asemănătoare unui șarpe. Există o caracteristică pe care trebuie să o prezinte o proteină. Structura aminoacizilor de pe ramuri ar trebui să fie cât mai scurtă posibil, cum ar fi glicina sau alanina, de exemplu. Acest tip de conformație secundară se numește foi beta pentru capacitatea lor de a lipi împreună pentru a forma o structură comună.
- Biologia se referă la al treilea tip de structură proteică ca fiind fragmente complexe, dispersate eterogen, dezordonate, care nu au stereoregularitate și sunt capabile să schimbe structura sub influența condițiilor externe.
Nu au fost identificate exemple de proteine care au în mod natural structură secundară.

Educatie tertiara
Aceasta este o conformație destul de complexă numită „globul”. Ce este această proteină? Structura sa se bazează pe structura secundară, totuși, se adaugă noi tipuri de interacțiuni între atomii grupelor, iar întreaga moleculă pare să se plieze, concentrându-se astfel asupra faptului că grupările hidrofile sunt direcționate în glob, iar cele hidrofobe. cele spre exterior.
Aceasta explică încărcarea moleculei de proteine în soluțiile coloidale de apă. Ce tipuri de interacțiuni sunt prezente aici?
- Legături de hidrogen – rămân neschimbate între aceleași părți ca și în structura secundară.
- interacțiuni – apar atunci când polipeptida este dizolvată în apă.
- Atractiile ionice se formează între grupuri încărcate diferit de resturi de aminoacizi (radicali).
- Interacțiuni covalente - se pot forma între anumite site-uri acide - molecule de cisteină, sau mai degrabă, cozile lor.
Astfel, compoziția și structura proteinelor cu structură terțiară pot fi descrise ca lanțuri polipeptidice pliate în globule care își păstrează și stabilizează conformația datorită diferitelor tipuri de interacțiuni chimice. Exemple de astfel de peptide: fosfoglicerat kenază, ARNt, alfa-keratina, fibroină de mătase și altele.
Structura cuaternară
Acesta este unul dintre cele mai complexe globule pe care le formează proteinele. Structura și funcțiile proteinelor de acest tip sunt foarte multifațetate și specifice.
Ce este această conformație? Acestea sunt mai multe (în unele cazuri zeci) lanțuri polipeptidice mari și mici care se formează independent unele de altele. Dar apoi, datorită acelorași interacțiuni pe care le-am considerat pentru structura terțiară, toate aceste peptide se răsucesc și se împletesc unele cu altele. În acest fel, se obțin globule conformaționale complexe, care pot conține atomi de metal, grupări lipidice și carbohidrați. Exemple de astfel de proteine: ADN polimeraza, învelișul proteic al virusului tutunului, hemoglobina și altele.

Toate structurile peptidice pe care le-am examinat au metode proprii de identificare în laborator, bazate pe capacități moderne de utilizare a cromatografiei, centrifugarei, microscopiei electronice și optice și a tehnologiilor informatice înalte.
Funcții îndeplinite
Structura și funcțiile proteinelor sunt strâns corelate între ele. Adică fiecare peptidă joacă un rol specific, unic și specific. Există, de asemenea, cei care sunt capabili să efectueze mai multe operații semnificative simultan într-o celulă vie. Cu toate acestea, este posibil să se exprime într-o formă generalizată principalele funcții ale moleculelor de proteine în organismele vii:
- Oferind mișcare. Organismele unicelulare sau organitele sau unele tipuri de celule sunt capabile de mișcare, contracție și mișcare. Acest lucru este asigurat de proteinele care alcătuiesc structura aparatului lor motor: cilii, flageli și membrana citoplasmatică. Dacă vorbim despre celulele incapabile de mișcare, atunci proteinele pot contribui la contracția acestora (miozina musculară).
- Funcția de nutriție sau de rezervă. Este acumularea de molecule de proteine în ouă, embrioni și semințele plantelor pentru a completa în continuare nutrienții lipsă. Când sunt defalcate, peptidele produc aminoacizi și substanțe biologic active care sunt necesare pentru dezvoltarea normală a organismelor vii.
- Funcția energetică. Pe lângă carbohidrați, proteinele pot oferi și putere organismului. Descompunerea a 1 g de peptidă eliberează 17,6 kJ de energie utilă sub formă de acid adenozin trifosforic (ATP), care este cheltuită pentru procesele vitale.
- Semnalizarea constă în monitorizarea atentă a proceselor în desfășurare și transmiterea semnalelor de la celule la țesuturi, de la acestea la organe, de la acestea din urmă către sisteme și așa mai departe. Un exemplu tipic este insulina, care fixează strict cantitatea de glucoză din sânge.
- Funcția receptorului. Se realizează prin modificarea conformației peptidei pe o parte a membranei și implicând celălalt capăt în restructurare. În același timp, semnalul și informațiile necesare sunt transmise. Cel mai adesea, astfel de proteine sunt încorporate în membranele citoplasmatice ale celulelor și exercită un control strict asupra tuturor substanțelor care trec prin ele. Ele oferă, de asemenea, informații despre modificările chimice și fizice din mediu.
- Funcția de transport a peptidelor. Este realizat de proteinele canal și proteinele transportoare. Rolul lor este evident - transportul moleculelor necesare în locuri cu concentrație scăzută din părți cu concentrație mare. Un exemplu tipic este transportul de oxigen și dioxid de carbon prin organe și țesuturi de către hemoglobina proteică. De asemenea, ei efectuează livrarea compușilor cu greutate moleculară mică prin membrana celulară în interior.
- Funcția structurală. Una dintre cele mai importante funcții îndeplinite de proteine. Structura tuturor celulelor și a organelelor lor este asigurată de peptide. Ei, ca un cadru, stabilesc forma și structura. În plus, îl susțin și îl modifică dacă este necesar. Prin urmare, pentru creștere și dezvoltare, toate organismele vii au nevoie de proteine în dieta lor. Astfel de peptide includ elastina, tubulina, colagenul, actina, keratina și altele.
- Funcția catalitică. Este realizat de enzime. Numeroase și variate, accelerează toate reacțiile chimice și biochimice din organism. Fără participarea lor, un măr obișnuit din stomac ar putea fi digerat în doar două zile, putrezind cel mai probabil în acest proces. Sub influența catalazei, peroxidazei și a altor enzime, acest proces are loc în două ore. În general, datorită acestui rol al proteinelor se realizează anabolismul și catabolismul, adică plastic și

Rol protector
Există mai multe tipuri de amenințări împotriva cărora proteinele sunt concepute pentru a proteja organismul.
În primul rând, reactivi traumatici, gaze, molecule, substanțe cu diferite spectre de acțiune. Peptidele sunt capabile să interacționeze chimic cu ele, transformându-le într-o formă inofensivă sau pur și simplu neutralizându-le.
În al doilea rând, amenințarea fizică a rănilor - dacă fibrinogenul proteic nu este transformat în fibrină la locul leziunii în timp, atunci sângele nu se va coagula, ceea ce înseamnă că nu va apărea blocarea. Apoi, dimpotrivă, veți avea nevoie de plasmină peptidică, care poate dizolva cheagul și poate restabili permeabilitatea vasului.
În al treilea rând, o amenințare la adresa imunității. Structura și semnificația proteinelor care formează apărarea imună sunt extrem de importante. Anticorpi, imunoglobuline, interferoni - toate acestea sunt elemente importante și semnificative ale sistemului limfatic și imunitar uman. Orice particulă străină, moleculă dăunătoare, parte moartă a unei celule sau o structură întreagă este supusă examinării imediate de către compusul peptidic. De aceea, o persoană se poate proteja în mod independent, fără ajutorul medicamentelor, de infecții și viruși simpli.
Proprietăți fizice
Structura unei proteine celulare este foarte specifică și depinde de funcția îndeplinită. Dar proprietățile fizice ale tuturor peptidelor sunt similare și se reduc la următoarele caracteristici.
- Greutatea moleculei este de până la 1.000.000 de daltoni.
- Sistemele coloidale se formează într-o soluție apoasă. Acolo structura capătă o încărcătură care poate varia în funcție de aciditatea mediului.
- Când sunt expuși la condiții dure (iradiere, acid sau alcali, temperatură etc.) sunt capabili să treacă la alte niveluri de conformații, adică denaturare. Acest proces este ireversibil în 90% din cazuri. Cu toate acestea, există și o schimbare inversă - renaturare.
Acestea sunt principalele proprietăți ale caracteristicilor fizice ale peptidelor.